版权声明:
2018 中国力学学会 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Atomic force microscopy (AFM) based force spectroscopy is a high-sensitive mechanical detection method. With unprecedented accuracy, this method enables the characterization of a wide range of biological and synthetic bio-interfaces, ranging from tissues, cells, membranes, proteins, nucleic acids to functional materials. Besides the possibility of high resolution imaging of biological samples from the cellular level to the molecular level, AFM-based force spectroscopy allows their mechanical, chemical, conductive or electrostatic, and biological properties to be probed, and helps addressing fundamental challenges that cannot be addressed with other techniques. In this review, we summarize basic and advanced force spectroscopy approaches in micro biomechanics and evaluate their unique advantages and limitations.
Keywords:atomic force microscopy
;
biomechanics
;
molecular force spectroscopy
;
intermolecular interactions
图6 原子力显微镜力谱技术作用于核酸的各类实验设计. (a)外力诱导下的DNA双螺旋熔融(Krautbauer et al. 2002b), (b)单链DNA与表面的摩擦力测试(Kühner et al. 2006), (c)将RNA链从病毒中直接提取的力学实验(Liu et al. 2010b), (d)外力诱导的DNA双螺旋熔融, 及其在SSB蛋白辅助下的复原(Liu et al. 2010a)
DNA的力曲线具有指纹特征, 只要是双螺旋DNA在拉伸条件下都会存在B-OS 转变平台.而许多DNA结合分子与DNA作用后会引起该转变区域指纹谱的变化, 因此该指纹谱是研究DNA结合试剂(如蛋白质和药物)与DNA相互作用的指示标志(Krautbauer et al.2002b, Krautbauer et al. 2000).原子力显微镜力谱实验中, 探针相对于样品的移动可以不限于在同一点的位置上仅沿\(Z\)轴上下移动, 在系统程序设定的路径下其活动范围可以是整个三维空间. 图6(b)描述了力谱技术用于研究不同聚合物样品(单链DNA, 聚烯丙胺)与云母表面的摩擦力测试(Kühner et al. 2006). 在实验过程中, 探针末端按照指定的路线接近或远离基底表面, 聚合物样品将表现出与表面解离或在表面滑动的现象, 也有可能同时发生这两种现象.于是此时获得的力谱信息, 将能够得到在设定的实验条件下聚合物链样品的摩擦力强弱的信息. 该研究表明, 即便两种聚合物链样品与同一表面具有类似大小的结合力, 它们与表面的摩擦力所反映的面内移动能力的差异仍然有可能具有十分显著的差异.
在涉及到DNA的复杂力谱力学操纵实验中, 可以利用互补单链DNA之间会形成DNA双螺旋的特点, 将单个目标分子剪切并黏贴到特定的位置, 与传统方法相比将定位精度显著提高到纳米量级(Heucke et al. 2014).此外, 通过使用表面电位循环技术结合力学操纵, 可以将原子力显微镜尖端的单分子DNA沉积在裸金电极上(Erdmann et al. 2010). 应用抗体和抗原肽标签, 单分子黏贴技术可以处理单一蛋白质.蛋白质--DNA复合物可以排列成复杂的样式, 同时保持蛋白质部分的功能.这种用于蛋白质排列的定向组装的单分子方法, 使得基于蛋白质组分的受控系统构建成为可能(Strackharn et al. 2012).经单分子力谱剪切进行的表面组装, 还可以用纳米级精度实时监测该过程.通过全内反射荧光显微镜和原子力显微镜的组合, 可以观察到纳米尺度图案沉积的单个荧光团(Kufer et al. 2009).虽然图案的光学分辨率远低于显微镜, 但可以通过将结合位点定位, 并检测荧光团的光漂白来识别各个染料. 通过这种方法的组合, 可以随意布置单独的染料标记的生物分子以用于特定功能, 例如偶联的荧光团系统或定制的酶级联, 同时辅以纳米级精度的实时监测.
除此以外, 通过基于力谱技术的原子力显微镜设备, 可以精确控制悬臂梁和探针在样品上移动轨迹和所施加的作用力.再加上可以选择具有不同物理性质的探针, 如镀有铂、金、钛等导电层或者镀有类金刚石耐磨层, 辅以施加在探针或样品上的电压, 还可以进行刻蚀和操纵实验等(Pires et al. 2010). 既可以应用于如图14所示的金属或半导体基底, 也可以用于生物样品表面的刻蚀和特定结构刻画(Liu et al. 2004, Paul et al.2011). 对基于原子力显微镜的力学操纵和刻蚀技术, Garcia等(2014)的综述文献中进行了详细报道.
图15 将各种小分子修饰于针尖后研究其与配对目标的相互作用. (a)葡萄糖(Puntheeranurak et al. 2007), (b) 抗癌多肽(Shan et al. 2012), (c) MFZ2-12分子(Wildling et al. 2012), (d)聚环氧乙烷(Sonnenberg et al. 2007b)
图19 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等.
Atomic force microscopy-based single-cell indentation: experimentation and finite element simulation. Proceedings of IEEE// International Conference on Intelligent Robots and Systems,
Analysis of the effect of LRP-1 silencing on the invasive potential of cancer cells by nanomechanical probing and adhesion force measurements using atomic force microscopy.
Molecular recognition force spectroscopy study of the dynamic interaction between aptamer GBI-10 and extracellular matrix protein tenascin-C on human glioblastoma cell.
Investigation of free fatty acid associated recombinant membrane receptor protein expression in HEK293 cells using raman spectroscopy, calcium imaging, and atomic force microscopy.
Extracting a single polyethylene oxide chain from a single crystal by a combination of atomic force microscopy imaging and single-molecule force spectroscopy: toward the investigation of molecular interactions in their condensed states.
Journal of the American Chemical Society, 133: 3226-3229.
Planned nanostructures of colloidal gold via self-assembly on hierarchically assembled organic bilayer template patterns with in-situ generated terminal amino functionality.
Park JW, Park AY, LeeS, Yu NK, Lee SH, Kaang BK.2010.
Detection of TRKB receptors distributed in cultured hippocampal neurons through bioconjugation between highly luminescent (quantum dot-neutravidin) and (biotinylated anti-TRKB antibody) on neurons by combined atomic force microscope and confocal laser scanning microscope.
Bioconjugate Chemistry, 21: 597-603.
[243]
Park PS, Sapra KT, Koli\'{n}skiM, FilipekS, PalczewskiK, Müller DJ.2007.
Stabilizing effect of Zn\(^{2 + }\) in native bovine rhodopsin.
Journal of Biological Chemistry, 282: 11377-11385.
[244]
Paul PC, Knoll AW, HolznerF, DespontM, DuerigU.2011.
Exploring the folding pattern of a polymer chain in a single crystal by combining single-molecule force spectroscopy and steered molecular dynamics simulations.
ThomaJ, BosshartP, PfreundschuhM, Müller D J. 2012. Out but not in: The large transmembrane b-barrel protein FhuA unfolds but cannot refold via b-hairpins. Structure, 20: 2185-2190.
Quantitative changes in the elasticity and adhesive properties of Escherichia coli ZK1056 prey cells during predation by Bdellovibrio bacteriovorus 109J.
Langmuir, 24: 8102-8110.
[317]
Walther KA, GräterF, DouganL, Badilla CL, Berne BJ, Fern\'{a}ndez JM.2007.
Signatures of hydrophobic collapse in extended proteins captured with force spectroscopy.
Proceedings of the National Academy of Sciences of the United States of America, 104: 7916-7921.
EMSA and single-molecule force spectroscopy study of interactions between bacillus subtilis single-stranded DNA-binding protein and single-stranded DNA.
Zhang YH, Liu CJ, Shi WQ, Wang ZQ, Dai LM, ZhangX.2007.
Direct measurements of the interaction between pyrene and graphite in aqueous media by single molecule force spectroscopy: Understanding the pi-pi interactions.
Harnessing the damping properties of materials for high-speed atomic force microscopy.
1
2016
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
Ligand-modulated parallel mechanical unfolding pathways of maltose-binding proteins.
1
2011
... 对于较大的蛋白质, 其经常表现出多个承受力单元, 使得蛋白质的去折叠不是"全或无"的过程, 而是显示出一系列的中间态, 比如绿色荧光蛋白(GFP) (Bertz et al. 2008, Mickler et al. 2007)、强化黄色荧光蛋白(EYFP) (Perez-Jimenez et al. 2006)、T4溶菌酶(T4 lysozyme) (Peng & Li 2008)、艾滋病毒1型受体蛋白CD4(Perez-Jimenez et al. 2014)、人体晶状蛋白(Garcia-Manyes et al. 2015)、麦芽糖结合蛋白(maltose binding protein) (Aggarwal et al. 2011)以及巨肌蛋白激酶(titin kinase)等(Puchner & Gaub 2009, Pernigo et al. 2010). ...
Direct observation of disulfide isomerization in a single protein.
0
2011
Detection of antigen-antibody binding events with the atomic force microscope.
1
1997
... 1994年Florin等通过牛血清白蛋白将亲和素分子连接到AFM 针尖, 在覆盖了生物素分子的琼脂糖珠上测量了单对生物素--亲和素之间的相互作用力(160 \(\pm\) 20) pN. Hinterdorfer等(1996)研究了蛋白抗体与抗原的相互作用力, 并改进了蛋白质的固定方法, 通过一段PEG高分子聚合物链将蛋白质与表面相连, 这样不仅增加了抗体抗原的空间自由度, 而且能从力--距离曲线图上将其特异性相互作用与针尖和基底的非特异性相互作用力区分开来, 成为目前固定蛋白质的常用方法.在覆盖有抗原分子(人血清白蛋白)的云母表面测量了单对抗体-抗原之间的相互作用力(244 \(\pm\) 22) pN.将提纯的蛋白质分子吸附到平整基底进行分子力测量的优点是消除了细胞表面(如细胞形貌, 细胞膜的柔软动态特性)对蛋白质分子的影响, 从而显著提高了信噪比, 并且可通过对蛋白质分子同时进行形貌成像和分子力测量来证明检测结果的准确性, 因而提高了实验结果的可信度(Wang H et al. 2008).通过将提纯的蛋白质分子吸附到基底, 研究人员测量了不同类型蛋白质分子之间的相互作用力, 如铁蛋白(ferritin) (Allen et al. 1997)、钙黏蛋白(cadherin)(Baumgartner et al. 2000)、路易斯寡糖(sLeX)-选择素(selectin)(Zhang X et al. 2004)、Strep-Tactin与绿色荧光蛋白等(Baumann et al. 2016). ...
Unfolding individual als5p adhesion proteins on live cells.
0
2009
Force-induced formation and propagation of adhesion nanodomains in living fungal cells.
Fundamental high-speed limits in single-molecule single-cell and nanoscale force spectroscopies.
1
2016
... 结合力谱技术的原子力显微镜成像模式在扫描时会记录数十万条力曲线数据, 用以反映机械性能或样品间的相互作用.因此缩短单条力曲线数据的记录时间, 能够减少图像获取时间并动态记录生物学的过程.然而有些配体--受体键合或者化学相互作用在非常快的外力作用下, 是远离平衡态的破坏状态, 此时想要推导其平衡态的力学行为是比较困难的(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).因为在高加载速率下, 必须考虑悬臂梁的流体动力学阻力和物理限制(Amo & Garcia 2016). 所以想要反映这类目标接近平衡态时的相互作用, 需要缓慢地施加外力. 因此, 似乎有可能需要开发能够在非常大的范围选择加载速率的方法, 例如结合当前的超稳定AFM方向(King et al. 2009, Bull et al. 2014, Weafer et al. 2012)与超高速AFM方向(Dong et al. 2011, Rico et al. 2013). ...
High-speed AFM and applications to biomolecular systems.
1
2013
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
Tentative identification of a resilin gene in Drosophila melanogaster.
1
2001
... 弹性蛋白质在生物力学系统中是一个重要的功能基本单元, 因为它们具有理想的弹性力学强度和抗性. 弹性蛋白质是天然黏合剂, 可以作为细胞黏附和肌肉蛋白质弹性的基础, 也可以作为具有极好机械性能的结构材料.随着对蛋白质力学性质了解的不断深入, 采用蛋白质人工构建新型材料的研究有了重要的进展(Guerette et al. 1996, Smith et al. 1999, Ardell & Andersen 2001, Becker et al. 2003, Elvin et al. 2005, Lyons et al. 2007).例如人们已经能够利用已知的蛛丝蛋白质的结构合成具有非常好的延展性和强度的人工材料(Wong et al.2012), 这种方法也为制造下一代力学稳定的生物材料指明了方向(Heim et al. 2009). ...
Nanoscale biophysical properties of the cell surface galactosaminogalactan from the fungal pathogen aspergillus fumigatus.
1
2015
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
Single-cell force spectroscopy of probiotic bacteria.
Molecular nanosprings in spider capture-silk threads.
1
2003
... 弹性蛋白质在生物力学系统中是一个重要的功能基本单元, 因为它们具有理想的弹性力学强度和抗性. 弹性蛋白质是天然黏合剂, 可以作为细胞黏附和肌肉蛋白质弹性的基础, 也可以作为具有极好机械性能的结构材料.随着对蛋白质力学性质了解的不断深入, 采用蛋白质人工构建新型材料的研究有了重要的进展(Guerette et al. 1996, Smith et al. 1999, Ardell & Andersen 2001, Becker et al. 2003, Elvin et al. 2005, Lyons et al. 2007).例如人们已经能够利用已知的蛛丝蛋白质的结构合成具有非常好的延展性和强度的人工材料(Wong et al.2012), 这种方法也为制造下一代力学稳定的生物材料指明了方向(Heim et al. 2009). ...
Models for the specific adhesion of cells to cells.
2
1978
... 测量断键力随外力加载速率的变化, 便可获得动力学力谱(dynamic force spectroscopy), 并计算出在不施加外力的平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\)、平均寿命\(t_{0}\)、活化态到结合态的距离\(x_{\rm u}\)等动力学参数, 以及复合物在解离过程中要跨越几个势垒, 是否存在复合物解离的中间体等(Lee et al. 2007, Strunz et al. 2000).单分子力谱是单分子水平研究生物分子复合物解离途径和解离动力学过程的有效方法.根据Bell-Evans模型, 施加复合物的断键方向上的力会减小复合物解离的活化能, 增大其解离速率常数, 单对分子复合物的断键力\(F\)与ln\(r\)\((r\)为外力加载速率)之间存在线性关系(Bell 1978, Evans & Ritchie 1997). 此外, 单分子力谱也可用来估计针尖上的配体与基底上受体之间的结合速率常数\(k_{\rm on}\)(Baumgartner et al. 2000).测量不同作用力下单分子相互作用的寿命, 就可获得势垒的位置与高度, 结合平衡态的热力学研究, 就可较完整地描绘出能量形貌图(Merkel et al. 1999). ...
... 膜蛋白去折叠过程会导致力曲线上产生很多锯齿状的突变峰, 每个突变峰均对应膜蛋白一个片段的去折叠过程.基于Bell-Evans模型推导出了蛋白质去折叠所需力与拉伸速率之间的关系, 揭示了蛋白质力学强度的动力学特性. 当拉伸速率较低时, 去折叠力将正比于拉伸速率, 对于较大的拉伸速率, 去折叠力与拉伸速率成指数关系(Bell 1978, Evans & Ritchie 1997). 通常利用虫链模型(worm-like chain)对力曲线上的各个突变峰进行拟合, 在虫链模型WLC拟合所得到的4个参数中, 每个结构域的能量势垒\(\DeltaG_{0}^\ddag \)的大小反映了跨越该能量势垒的难度, 此值越大越需要更大的作用力去克服, 可以反映结构域的稳定性; 活化态到结合态的距离\(x_{\rm u}\)反映了结构域能够容纳的构象变化, 此值越大则代表在去折叠过程中结构域可能出现的构象形态越多; 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\), 此值越大说明在自然条件下发生解离的速率越大, 结构域不稳定性越高; 等效弹性系数\(\kappa \)反映了结构域的机械力学参数, 此值越大表示结构域的机械硬度越强.通过上述参数可以描绘出去折叠动态过程的能量图谱(Janovjak et al. 2004, Kuo et al. 2010, Schoeler et al. 2015).也有研究表明这一能量图谱的表面粗糙而非平滑(Janovjak et al. 2007).近年来随着模型的优化已经有高度集成化的分析软件(Lamour et al. 2014), 同时随着设备性能的提升, 已经逐渐展开了这一模型在高速拉伸下的分析(Junker & Rief 2010, Berkovich et al. 2012). ...
Discrete interactions in cell adhesion measured by single-molecule force spectroscopy.
Relaxation of loaded ESCRT-III spiral springs drives membrane deformation.
1
2015
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
Protein misfolding functional amyloid and human disease.
Nano-scale dynamic recognition imaging on vascularendothelial cells.
1
2007
... 此方法的一般步骤是将一个可伸缩的线性分子连接于探针, 再将需固定的分子连接于线性分子, 这种方法使被连接的分子具有一定的空间自由度(Chtcheglova et al. 2007). 针尖化学修饰的方法有多种, 其中比较常用的修饰方法是商品化的硅或氮化硅探针经化学处理, 如食人鱼洗液(98% H\(_{2}\)SO\(_{4}\):30%H\(_{2}\)O\(_{2}\)体积比7:3)清洗, 使其表面暴露出大量硅羟基(---SiOH), 再对探针表面硅羟基进一步硅烷化、醚化或氢硅化等, 或者直接将镀金针尖进行巯基化处理. 其中, 探针的硅烷化基于探针表面活化的硅羟基与硅烷化试剂反应, 如3 -氨基丙基三乙氧基硅烷(3-aminopropyltriethoxysilane, APTES)、3 -氨基丙基二甲基甲氧硅烷(3-aminopropyldimethylmethoxysilane, APS)等, 在针尖表面形成含有所需化学基团的有机单分子层(Xue et al. 2014).探针的醚化是利用醇类(通常是乙醇胺)与探针表面活化的羟基(---OH)反应, 形成末端含有氨基(---NH\(_{2})\)的有机单分子层(Wildling et al. 2011).在探针的氢硅化中, 首先对探针表面用1%的氢氟酸(HF)处理形成氢化的硅表面, 然后利用氢化表面与烯烃加成反应, 通过Si---C共价键将有机分子连接到针尖表面(Buriak 2002).镀金层探针的化学修饰方法, 是镀金探针表面与烷硫醇的巯基通过金硫键之间的强非特异性相互作用, 形成稳定的自组装单分子层(Wang et al. 2004). ...
Observing growth steps of collagen self-assembly by time-lapse high-resolution atomic force microscopy.
1
2006
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Resolving single-molecule assembled patterns with superresolution blink-microscopy.
1
2010
... (5)原子力显微镜只能获得样品表面信息, 难以获得细胞或菌体等生物体内部分子的特异性信息, 因此需要与其他技术联合使用, 例如激光扫描共聚焦显微镜、荧光显微镜、透射电子显微镜、流式细胞仪及分子动力学模拟等(Te Riet et al. 2014, Ramanathan et al. 2015, Franz & Müller 2005 ,Cordes et al. 2010, Monserrate et al. 2014).这将成为原子力显微镜力谱技术的发展趋势.选择时光学显微镜的分辨率应尽可能高, 以便能够更好地将光学图像与不受衍射极限限制的AFM图像相关联. ...
Single-cell elastography: Probing for disease with the atomic force microscope.
1
2003
... 赫兹模型成立的两个前提条件是压入物呈抛物线形和样品厚度足够大(相对于针尖压细胞的深度), 同时要求样品表面连续且光滑, 实验过程中样品形变量相对较小(Kuznetsova et al. 2007, Costa 2003).当探针为球形时, 赫兹模型为 \[ f_{\rm sphere} =\dfrac{4}{3}\dfrac{E}{1 - v^2}\sqrt R \delta ^{3 / 2} \ \ (7)\] ...
Nanomechanical analysis of cells from cancer patients.
Weakly bound water molecules shorten single-stranded DNA.
1
2006
... 生物大分子在生理条件下(酸碱度近中性的室温稀盐水溶液)往往会形成规整、精确的超分子结构, 比如双链DNA 以双螺旋的形式存在, 蛋白质则经由\(\alpha \)-螺旋、\(\beta\)-折叠等结构完成组装. 为了研究生物大分子与水的相互作用, Cui等利用单分子力谱迫使高分子单链进入到不良溶剂中, 开展了生物大分子在水溶液与非极性溶剂中的对照研究(Cui et al. 2006, 2007). 双链DNA 在受到外力拉伸时, 可在约65 pN处观察到一个长平台, 对应于从B型双链DNA到其过度拉伸态的结构转变. 然而实验中发现, 在不同溶剂中力曲线存在着明显的变化.如果将水溶液替换为非极性溶剂(如二乙苯等), 不仅无法观测到指纹特征, 而且所得力曲线与单链DNA的一致.这一结果表明双链DNA在非极性溶剂中将变性为单链结构.在实验结果的基础上, 提出ssDNA与水分子之间的弱氢键可能是双链DNA稳定存在于水中的前提条件.原子力显微镜也被广泛应用与除DNA之外的许多小分子的力谱研究, 甚至包括了金颗粒(Xue et al. 2014)、纳米线(Marszalek et al. 2000)等非生物样品, 如图7(b)所示.早期的研究应用实例包括应用力谱分析技术在单分子水平上鉴定多糖混合物的组分(Marszalek et al.2001), 观察黄原胶分子与基底的结合力(Li et al. 1999a), 研究蚕丝素分子的纳米力学特性(Zhang et al. 2000), 观察肝素分子在外力作用下的力学性质(Marszalek et al. 2003), Fern\'{a}ndez课题组对泛素分子力学性质、及其与水分子相互作用的一系列研究等(Carrion-Vazquez et al.2003, Schlierf & Fern\'{a}ndez 2004, Brujić et al. 2006, Li et al. 2010). ...
Double-stranded DNA dissociates into single strands when dragged into a poor solvent.
1
2007
... 生物大分子在生理条件下(酸碱度近中性的室温稀盐水溶液)往往会形成规整、精确的超分子结构, 比如双链DNA 以双螺旋的形式存在, 蛋白质则经由\(\alpha \)-螺旋、\(\beta\)-折叠等结构完成组装. 为了研究生物大分子与水的相互作用, Cui等利用单分子力谱迫使高分子单链进入到不良溶剂中, 开展了生物大分子在水溶液与非极性溶剂中的对照研究(Cui et al. 2006, 2007). 双链DNA 在受到外力拉伸时, 可在约65 pN处观察到一个长平台, 对应于从B型双链DNA到其过度拉伸态的结构转变. 然而实验中发现, 在不同溶剂中力曲线存在着明显的变化.如果将水溶液替换为非极性溶剂(如二乙苯等), 不仅无法观测到指纹特征, 而且所得力曲线与单链DNA的一致.这一结果表明双链DNA在非极性溶剂中将变性为单链结构.在实验结果的基础上, 提出ssDNA与水分子之间的弱氢键可能是双链DNA稳定存在于水中的前提条件.原子力显微镜也被广泛应用与除DNA之外的许多小分子的力谱研究, 甚至包括了金颗粒(Xue et al. 2014)、纳米线(Marszalek et al. 2000)等非生物样品, 如图7(b)所示.早期的研究应用实例包括应用力谱分析技术在单分子水平上鉴定多糖混合物的组分(Marszalek et al.2001), 观察黄原胶分子与基底的结合力(Li et al. 1999a), 研究蚕丝素分子的纳米力学特性(Zhang et al. 2000), 观察肝素分子在外力作用下的力学性质(Marszalek et al. 2003), Fern\'{a}ndez课题组对泛素分子力学性质、及其与水分子相互作用的一系列研究等(Carrion-Vazquez et al.2003, Schlierf & Fern\'{a}ndez 2004, Brujić et al. 2006, Li et al. 2010). ...
Vertical collapse of a cytolysin prepore moves its transmembrane beta-hairpins to the membrane.
1
2004
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Chemical force microscopy of single live cells.
1
2007
... 与本文第4章的内容类似, 当使用化学基团、配体、受体或病毒功能化的探针进行测量时, 探针会与样品发生特异性相互作用.如果此时使用力谱技术同时结合成像功能, 则能够将这些相互作用的位点映射到样品的形貌图之上.这一技术类型的尝试, 早在21世纪初就已经开始.应用的实例包括探索化学基团、多糖或蛋白质与细胞表面的相互作用结合形貌的研究(Grandbois et al.2000, Dague et al. 2007, Hinterdorfer & Dufrêne2006)等. ...
One \(\beta \) hairpin follows the other exploring refolding pathways and kinetics of the transmembrane b-barrel protein OmpG.
1
2011
... 通过力谱技术对单个膜蛋白的力学操纵, 可以观察其重新折叠回到膜中的动力学过程.液相中能够直接折叠回到膜中的一般是只有部分去折叠的小蛋白质, 例如紫膜中的视紫红质蛋白质和OmpG蛋白质. 图13显示了将膜内的视紫红质蛋白质拉伸后, 使其部分去折叠①, 再将其按压至膜层表面②, 之后再次拉伸至只有最后一个结构域仍镶嵌于膜内③, 再次按压回表面④, 最后将其完全从膜内拉出⑦的复杂力谱实验.通过对视紫红质蛋白质的精确力学操纵, 发现部分去折叠的小型膜蛋白按压于膜层表面时, 其可以直接再次折叠回到膜内(Kessler et al. 2006).类似的实验结果也被发现于跨膜\(\beta \)折叠蛋白质OmpG中(Damaghi et al. 2011). ...
Binding strength between cell adhesion proto glycols measured by atomic force microscope.
On the measurements of rigidity modulus of soft materials in nanoindentation experiments at small depth.
1
2012
... (3)采用不同的模型测量细胞的杨氏模量得到的结果差别较大, 因此, 如何统一测量标准是需要解决的实际问题(Moreno-Flores et al. 2010, Guz et al. 2014, Dokukin & Sokolov 2012). ...
Determination of protein structural flexibility by microsecond force spectroscopy.
1
2009
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
A nanomechanical interface to rapid single-molecule interactions.
1
2011
... 结合力谱技术的原子力显微镜成像模式在扫描时会记录数十万条力曲线数据, 用以反映机械性能或样品间的相互作用.因此缩短单条力曲线数据的记录时间, 能够减少图像获取时间并动态记录生物学的过程.然而有些配体--受体键合或者化学相互作用在非常快的外力作用下, 是远离平衡态的破坏状态, 此时想要推导其平衡态的力学行为是比较困难的(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).因为在高加载速率下, 必须考虑悬臂梁的流体动力学阻力和物理限制(Amo & Garcia 2016). 所以想要反映这类目标接近平衡态时的相互作用, 需要缓慢地施加外力. 因此, 似乎有可能需要开发能够在非常大的范围选择加载速率的方法, 例如结合当前的超稳定AFM方向(King et al. 2009, Bull et al. 2014, Weafer et al. 2012)与超高速AFM方向(Dong et al. 2011, Rico et al. 2013). ...
Direct measurement of colloidal forces using an atomic force microscope.
1
1991
... 自原子力显微镜的早期应用以来, 力--距离曲线就已经被用于测量界面的机械性能, 并量化地测量范德华相互作用、疏水和亲水的性质, 以及离子层电荷和静电双层相互作用力等(Frisbie et al. 1994, Ducker et al. 1991). 在进针过程中, 原子力显微镜的探针与样品在指定位置相互作用, 直到达到指定的最大压力或者指定的最大位移. 力曲线趋近曲线部分的分析, 特别是描述压痕的区域, 可以定量的描述样品的变形、弹性和能量耗散的性质.力曲线回退曲线部分的分析, 能够描述探针与样品之间的黏附力.作用在探针和样品上的分子间、和分子内作用力, 一般取决于它们的物理化学性质和缓冲溶液(Butt et al. 2005). ...
Theory analysis and interpretation of single-molecule force spectroscopy experiments.
2
2008
... 随着研究领域的深入, 人们意识到测量不同分离速度(即加载速率)的断裂力提供了关于结合键的动力学性质的信息(Moy et al.1994).结合键的动力学模型分析可以用于估计配体--受体键的热力学和动力学性质(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).这些性质包括跨越折叠状态的能量势垒\(\Delta G_{0}^\ddag \); 活化态到结合态的距离\(x_{\rm u}\); 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\); 此外结合键的寿命也可以通过使键处于低恒定力并等待键的断裂来测量(Woodside & Block 2014, Perez-Jimenez et al. 2011).此恒定作用力可以通过原子力显微镜的反馈系统以力钳模式施加(Oberhauser et al.2001, Stahl et al. 2009), 也可以利用天然的细胞膜栓系Tether来实现(Krieg et al. 2008b). ...
... 结合力谱技术的原子力显微镜成像模式在扫描时会记录数十万条力曲线数据, 用以反映机械性能或样品间的相互作用.因此缩短单条力曲线数据的记录时间, 能够减少图像获取时间并动态记录生物学的过程.然而有些配体--受体键合或者化学相互作用在非常快的外力作用下, 是远离平衡态的破坏状态, 此时想要推导其平衡态的力学行为是比较困难的(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).因为在高加载速率下, 必须考虑悬臂梁的流体动力学阻力和物理限制(Amo & Garcia 2016). 所以想要反映这类目标接近平衡态时的相互作用, 需要缓慢地施加外力. 因此, 似乎有可能需要开发能够在非常大的范围选择加载速率的方法, 例如结合当前的超稳定AFM方向(King et al. 2009, Bull et al. 2014, Weafer et al. 2012)与超高速AFM方向(Dong et al. 2011, Rico et al. 2013). ...
Multiparametric imaging of biological systems by force-distance curve-based AFM.
1
2013
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
The yeast Wsc1 cell surface sensor behaves like a nanospring in vivo.
Synthesis and properties of crosslinked recombinant pro-resilin.
1
2005
... 弹性蛋白质在生物力学系统中是一个重要的功能基本单元, 因为它们具有理想的弹性力学强度和抗性. 弹性蛋白质是天然黏合剂, 可以作为细胞黏附和肌肉蛋白质弹性的基础, 也可以作为具有极好机械性能的结构材料.随着对蛋白质力学性质了解的不断深入, 采用蛋白质人工构建新型材料的研究有了重要的进展(Guerette et al. 1996, Smith et al. 1999, Ardell & Andersen 2001, Becker et al. 2003, Elvin et al. 2005, Lyons et al. 2007).例如人们已经能够利用已知的蛛丝蛋白质的结构合成具有非常好的延展性和强度的人工材料(Wong et al.2012), 这种方法也为制造下一代力学稳定的生物材料指明了方向(Heim et al. 2009). ...
Electrically induced bonding of DNA to gold.
1
2010
... 在涉及到DNA的复杂力谱力学操纵实验中, 可以利用互补单链DNA之间会形成DNA双螺旋的特点, 将单个目标分子剪切并黏贴到特定的位置, 与传统方法相比将定位精度显著提高到纳米量级(Heucke et al. 2014).此外, 通过使用表面电位循环技术结合力学操纵, 可以将原子力显微镜尖端的单分子DNA沉积在裸金电极上(Erdmann et al. 2010). 应用抗体和抗原肽标签, 单分子黏贴技术可以处理单一蛋白质.蛋白质--DNA复合物可以排列成复杂的样式, 同时保持蛋白质部分的功能.这种用于蛋白质排列的定向组装的单分子方法, 使得基于蛋白质组分的受控系统构建成为可能(Strackharn et al. 2012).经单分子力谱剪切进行的表面组装, 还可以用纳米级精度实时监测该过程.通过全内反射荧光显微镜和原子力显微镜的组合, 可以观察到纳米尺度图案沉积的单个荧光团(Kufer et al. 2009).虽然图案的光学分辨率远低于显微镜, 但可以通过将结合位点定位, 并检测荧光团的光漂白来识别各个染料. 通过这种方法的组合, 可以随意布置单独的染料标记的生物分子以用于特定功能, 例如偶联的荧光团系统或定制的酶级联, 同时辅以纳米级精度的实时监测. ...
Forces and bond dynamics in cell adhesion.
2
2007
... 随着研究领域的深入, 人们意识到测量不同分离速度(即加载速率)的断裂力提供了关于结合键的动力学性质的信息(Moy et al.1994).结合键的动力学模型分析可以用于估计配体--受体键的热力学和动力学性质(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).这些性质包括跨越折叠状态的能量势垒\(\Delta G_{0}^\ddag \); 活化态到结合态的距离\(x_{\rm u}\); 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\); 此外结合键的寿命也可以通过使键处于低恒定力并等待键的断裂来测量(Woodside & Block 2014, Perez-Jimenez et al. 2011).此恒定作用力可以通过原子力显微镜的反馈系统以力钳模式施加(Oberhauser et al.2001, Stahl et al. 2009), 也可以利用天然的细胞膜栓系Tether来实现(Krieg et al. 2008b). ...
... 结合力谱技术的原子力显微镜成像模式在扫描时会记录数十万条力曲线数据, 用以反映机械性能或样品间的相互作用.因此缩短单条力曲线数据的记录时间, 能够减少图像获取时间并动态记录生物学的过程.然而有些配体--受体键合或者化学相互作用在非常快的外力作用下, 是远离平衡态的破坏状态, 此时想要推导其平衡态的力学行为是比较困难的(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).因为在高加载速率下, 必须考虑悬臂梁的流体动力学阻力和物理限制(Amo & Garcia 2016). 所以想要反映这类目标接近平衡态时的相互作用, 需要缓慢地施加外力. 因此, 似乎有可能需要开发能够在非常大的范围选择加载速率的方法, 例如结合当前的超稳定AFM方向(King et al. 2009, Bull et al. 2014, Weafer et al. 2012)与超高速AFM方向(Dong et al. 2011, Rico et al. 2013). ...
Dynamic strength of molecular adhesion bonds.
2
1997
... 测量断键力随外力加载速率的变化, 便可获得动力学力谱(dynamic force spectroscopy), 并计算出在不施加外力的平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\)、平均寿命\(t_{0}\)、活化态到结合态的距离\(x_{\rm u}\)等动力学参数, 以及复合物在解离过程中要跨越几个势垒, 是否存在复合物解离的中间体等(Lee et al. 2007, Strunz et al. 2000).单分子力谱是单分子水平研究生物分子复合物解离途径和解离动力学过程的有效方法.根据Bell-Evans模型, 施加复合物的断键方向上的力会减小复合物解离的活化能, 增大其解离速率常数, 单对分子复合物的断键力\(F\)与ln\(r\)\((r\)为外力加载速率)之间存在线性关系(Bell 1978, Evans & Ritchie 1997). 此外, 单分子力谱也可用来估计针尖上的配体与基底上受体之间的结合速率常数\(k_{\rm on}\)(Baumgartner et al. 2000).测量不同作用力下单分子相互作用的寿命, 就可获得势垒的位置与高度, 结合平衡态的热力学研究, 就可较完整地描绘出能量形貌图(Merkel et al. 1999). ...
... 膜蛋白去折叠过程会导致力曲线上产生很多锯齿状的突变峰, 每个突变峰均对应膜蛋白一个片段的去折叠过程.基于Bell-Evans模型推导出了蛋白质去折叠所需力与拉伸速率之间的关系, 揭示了蛋白质力学强度的动力学特性. 当拉伸速率较低时, 去折叠力将正比于拉伸速率, 对于较大的拉伸速率, 去折叠力与拉伸速率成指数关系(Bell 1978, Evans & Ritchie 1997). 通常利用虫链模型(worm-like chain)对力曲线上的各个突变峰进行拟合, 在虫链模型WLC拟合所得到的4个参数中, 每个结构域的能量势垒\(\DeltaG_{0}^\ddag \)的大小反映了跨越该能量势垒的难度, 此值越大越需要更大的作用力去克服, 可以反映结构域的稳定性; 活化态到结合态的距离\(x_{\rm u}\)反映了结构域能够容纳的构象变化, 此值越大则代表在去折叠过程中结构域可能出现的构象形态越多; 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\), 此值越大说明在自然条件下发生解离的速率越大, 结构域不稳定性越高; 等效弹性系数\(\kappa \)反映了结构域的机械力学参数, 此值越大表示结构域的机械硬度越强.通过上述参数可以描绘出去折叠动态过程的能量图谱(Janovjak et al. 2004, Kuo et al. 2010, Schoeler et al. 2015).也有研究表明这一能量图谱的表面粗糙而非平滑(Janovjak et al. 2007).近年来随着模型的优化已经有高度集成化的分析软件(Lamour et al. 2014), 同时随着设备性能的提升, 已经逐渐展开了这一模型在高速拉伸下的分析(Junker & Rief 2010, Berkovich et al. 2012). ...
Quantification of surface tension and internal pressure generated by single mitotic cells.
The study of protein mechanics with the atomic force microscope.
1
1999
... 原子力显微镜力谱技术研究单个蛋白质分子去折叠过程. (a)肌联蛋白Titin在外力所用下的机械展开(Fisher et al. 1999); (b)将单个细菌视紫红质分子从紫膜表面提拉迟来, 并对比拉伸前后的细菌紫膜表面成像区别(Oesterhelt et al. 2000) ...
Adhesion forces between individual ligand-receptor pairs.
0
1994
Zinc-dependent mechanical properties of Staphylococcus aureus biofilm-forming surface protein SasG.
1
2016
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
Detection localization and conformational analysis of single polysaccharide molecules on live bacteria.
Cellular remodelling of individual collagen fibrils visualized by time-lapse AFM.
3
2007
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
... )、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
... 原子力显微镜用于各尺寸生物样品成像示例. (a)核酸(Pyne et al. 2014); (b)蛋白质(Seelert et al. 2000); (c)纤维(Stamov et al. 2015); (d)病毒(Kuznetsov & McPherson 2011); (e)细胞(Friedrichs et al. 2007); (f)秀丽线虫横截面, 扫描范围24 \(\mu \)m \(\times \) 24 \(\mu \)m \(\times\) 6um (NT-MDT公司) ...
Functional group imaging by chemical force microscopy.
1
1994
... 自原子力显微镜的早期应用以来, 力--距离曲线就已经被用于测量界面的机械性能, 并量化地测量范德华相互作用、疏水和亲水的性质, 以及离子层电荷和静电双层相互作用力等(Frisbie et al. 1994, Ducker et al. 1991). 在进针过程中, 原子力显微镜的探针与样品在指定位置相互作用, 直到达到指定的最大压力或者指定的最大位移. 力曲线趋近曲线部分的分析, 特别是描述压痕的区域, 可以定量的描述样品的变形、弹性和能量耗散的性质.力曲线回退曲线部分的分析, 能够描述探针与样品之间的黏附力.作用在探针和样品上的分子间、和分子内作用力, 一般取决于它们的物理化学性质和缓冲溶液(Butt et al. 2005). ...
Advanced scanning probe lithography.
1
2014
... 除此以外, 通过基于力谱技术的原子力显微镜设备, 可以精确控制悬臂梁和探针在样品上移动轨迹和所施加的作用力.再加上可以选择具有不同物理性质的探针, 如镀有铂、金、钛等导电层或者镀有类金刚石耐磨层, 辅以施加在探针或样品上的电压, 还可以进行刻蚀和操纵实验等(Pires et al. 2010). 既可以应用于如图14所示的金属或半导体基底, 也可以用于生物样品表面的刻蚀和特定结构刻画(Liu et al. 2004, Paul et al.2011). 对基于原子力显微镜的力学操纵和刻蚀技术, Garcia等(2014)的综述文献中进行了详细报道. ...
Force-activated reactivity switch in a bimolecular chemical reaction.
0
2009
Single molecule force spectroscopy predicts a misfolded domain-swapped conformation in human \(\gamma \)D-crystallin.
1
2015
... 对于较大的蛋白质, 其经常表现出多个承受力单元, 使得蛋白质的去折叠不是"全或无"的过程, 而是显示出一系列的中间态, 比如绿色荧光蛋白(GFP) (Bertz et al. 2008, Mickler et al. 2007)、强化黄色荧光蛋白(EYFP) (Perez-Jimenez et al. 2006)、T4溶菌酶(T4 lysozyme) (Peng & Li 2008)、艾滋病毒1型受体蛋白CD4(Perez-Jimenez et al. 2014)、人体晶状蛋白(Garcia-Manyes et al. 2015)、麦芽糖结合蛋白(maltose binding protein) (Aggarwal et al. 2011)以及巨肌蛋白激酶(titin kinase)等(Puchner & Gaub 2009, Pernigo et al. 2010). ...
Switching the mechanics of dsDNA by Cu salicylic aldehyde complexation.
1
2009
... 核酸本身的双螺旋结构即可作为配对目标的一种典型研究.通过将单链DNA修饰在原子力显微镜的针尖, 对其与互补DNA单链的相互作用展开了一系列研究(Moy et al. 1995).检测出双链DNA的解链作用力大小为每10碱基对10 \(\sim\) 20 pN(Krautbauer et al. 2003).通过对两个30碱基的寡核苷酸双链体组成的分子力平衡的实验, 给出了一种能够以设定的外力和加载速率模拟力诱导下的断裂模型.并用DNA双链两端双键的断裂位点进行了预测(Neuert et al. 2007).核酸正在越来越多地作为纳米尺度结构的可编程构建块.通过特定DNA碱基对识别进行自组装, 可以形成各种特异性结构.这些结构的稳定化可以通过与取代碱基的金属配位键来实现.在携带水杨醛核苷的双链DNA在铜金属络合时的力学转换研究中, 发现断键力增大了两倍. 动力学分析表明在外力加载速率较高时, 此生物分子杂交体的力学性质主要受局域配位键的影响, 而在外力加载速率较低时, 目标的稳定性主要受碱基对相互结合的影响(Gaub et al. 2009). ...
Determination of the elastic moduli of thin samples and adherent cells using conical atomic force microscope tips.
... 与本文第4章的内容类似, 当使用化学基团、配体、受体或病毒功能化的探针进行测量时, 探针会与样品发生特异性相互作用.如果此时使用力谱技术同时结合成像功能, 则能够将这些相互作用的位点映射到样品的形貌图之上.这一技术类型的尝试, 早在21世纪初就已经开始.应用的实例包括探索化学基团、多糖或蛋白质与细胞表面的相互作用结合形貌的研究(Grandbois et al.2000, Dague et al. 2007, Hinterdorfer & Dufrêne2006)等. ...
Studying early stages of fibronectin fibrillogenesis in living cells by atomic force microscopy.
1
2015
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Silk properties determined by gland-specific expression of a spider fibroin gene family.
1
1996
... 弹性蛋白质在生物力学系统中是一个重要的功能基本单元, 因为它们具有理想的弹性力学强度和抗性. 弹性蛋白质是天然黏合剂, 可以作为细胞黏附和肌肉蛋白质弹性的基础, 也可以作为具有极好机械性能的结构材料.随着对蛋白质力学性质了解的不断深入, 采用蛋白质人工构建新型材料的研究有了重要的进展(Guerette et al. 1996, Smith et al. 1999, Ardell & Andersen 2001, Becker et al. 2003, Elvin et al. 2005, Lyons et al. 2007).例如人们已经能够利用已知的蛛丝蛋白质的结构合成具有非常好的延展性和强度的人工材料(Wong et al.2012), 这种方法也为制造下一代力学稳定的生物材料指明了方向(Heim et al. 2009). ...
Measuring protein isoelectric points by AFM-based force spectroscopy using trace amounts of sample.
1
2016
... 这些早期的实验主要受到设备采样率和压电陶瓷性能的限制, 其每条力曲线都需要接近数秒钟的时间完成, 这就大大制约了能够采集的力曲线的数量.由于每个像素至少要有一条力曲线才能计算机械性能, 所以早期的功能探针力谱结合成像技术结果往往像素点都比较少, 且实验时间比较长. 最近五年来, 随着设备性能的提升、新模式的推广, 以及探针相对样品运动轨迹的改良, 商品化设备的力曲线采集频率已经可以提升到2 kHz左右, 使得配对样品的高通量表征成为了现实(见图21).这方面的应用实例包括: 生物膜和膜蛋白的静电性质(Medalsy & Müller 2013, Guo et al. 2016, Pfreundschuh et al. 2013), 被细菌排出的噬菌体(Alsteens et al. 2013a), 结合动物细胞表面受体的包膜病毒(Alsteens et al. 2017b), 结合人G蛋白偶联受体的配体(Alsteens et al. 2015), 两个配体与相同受体的结合(Pfreundschuh et al. 2015), 以及大型蛋白质复合物上配体结合事件的高清成像等(Pfreundschuh et al. 2014, Kim et al. 2015). ...
If cell mechanics can be described by elastic modulus: study of different models and probes used in indentation experiments.
1
2014
... (3)采用不同的模型测量细胞的杨氏模量得到的结果差别较大, 因此, 如何统一测量标准是需要解决的实际问题(Moreno-Flores et al. 2010, Guz et al. 2014, Dokukin & Sokolov 2012). ...
High-speed atomic force microscopy.
1
2006
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
Direct observation of the reversible two-state unfolding and refolding of an ab protein by single-molecule atomic force microscopy.
Mechanically tightening a protein slipknot into a trefoil knot.
0
2014
Imaging viscoelastic properties of live cells by AFM: Power-law rheology on the nanoscale.
1
2015
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
Spider silk: from soluble protein to extraordinary fiber.
1
2009
... 弹性蛋白质在生物力学系统中是一个重要的功能基本单元, 因为它们具有理想的弹性力学强度和抗性. 弹性蛋白质是天然黏合剂, 可以作为细胞黏附和肌肉蛋白质弹性的基础, 也可以作为具有极好机械性能的结构材料.随着对蛋白质力学性质了解的不断深入, 采用蛋白质人工构建新型材料的研究有了重要的进展(Guerette et al. 1996, Smith et al. 1999, Ardell & Andersen 2001, Becker et al. 2003, Elvin et al. 2005, Lyons et al. 2007).例如人们已经能够利用已知的蛛丝蛋白质的结构合成具有非常好的延展性和强度的人工材料(Wong et al.2012), 这种方法也为制造下一代力学稳定的生物材料指明了方向(Heim et al. 2009). ...
... 力谱技术为表征生物系统的机械特性提供了有力的工具, 但是理论推导和实验结果均表明, 这些系统的机械响应是取决于施加力的加载速率(loading rate) (Janmey et al.2007, Hinterdorfer & Dufrêne 2006).加载速率是指施加于被测物体的作用力随时间的变化率, 以某一恒定速率拉回探针时测定的分子间解离力, 只是一个连续的分子间作用力谱中的一个点.由于系统的机械响应与作用力的加载速率是非线性关系, 并且随着生物系统的不同而变化, 因此不建议仅以单一的加载速率进行力谱实验.为了更好地了解生物系统的机械性能, 一般需要在较宽范围的加载速率下对该响应进行表征(Krieg et al. 2014, Medalsy & Müller 2013, Herruzo et al. 2014). ...
Glyphosate-induced stiffening of HaCaT keratinocytes, a peak force tapping study on living cells.
1
2012
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
Placing individual molecules in the center of nanoapertures.
1
2014
... 在涉及到DNA的复杂力谱力学操纵实验中, 可以利用互补单链DNA之间会形成DNA双螺旋的特点, 将单个目标分子剪切并黏贴到特定的位置, 与传统方法相比将定位精度显著提高到纳米量级(Heucke et al. 2014).此外, 通过使用表面电位循环技术结合力学操纵, 可以将原子力显微镜尖端的单分子DNA沉积在裸金电极上(Erdmann et al. 2010). 应用抗体和抗原肽标签, 单分子黏贴技术可以处理单一蛋白质.蛋白质--DNA复合物可以排列成复杂的样式, 同时保持蛋白质部分的功能.这种用于蛋白质排列的定向组装的单分子方法, 使得基于蛋白质组分的受控系统构建成为可能(Strackharn et al. 2012).经单分子力谱剪切进行的表面组装, 还可以用纳米级精度实时监测该过程.通过全内反射荧光显微镜和原子力显微镜的组合, 可以观察到纳米尺度图案沉积的单个荧光团(Kufer et al. 2009).虽然图案的光学分辨率远低于显微镜, 但可以通过将结合位点定位, 并检测荧光团的光漂白来识别各个染料. 通过这种方法的组合, 可以随意布置单独的染料标记的生物分子以用于特定功能, 例如偶联的荧光团系统或定制的酶级联, 同时辅以纳米级精度的实时监测. ...
Detection and localization of individual antibody-antigen recognition events by atomic force microscopy.
Immunoactive two-dimensional self-assembly of monoclonal antibodies in aqueous solution revealed by atomic force microscopy.
1
2014
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Beyond the helix pitch: Direct visualization of native DNA in aqueous solution.
1
2013
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Atomic force microscopy detects differences in the surface brush of normal and cancerous cells.
0
2009
Basic rheology for biologists.
1
2007
... 力谱技术为表征生物系统的机械特性提供了有力的工具, 但是理论推导和实验结果均表明, 这些系统的机械响应是取决于施加力的加载速率(loading rate) (Janmey et al.2007, Hinterdorfer & Dufrêne 2006).加载速率是指施加于被测物体的作用力随时间的变化率, 以某一恒定速率拉回探针时测定的分子间解离力, 只是一个连续的分子间作用力谱中的一个点.由于系统的机械响应与作用力的加载速率是非线性关系, 并且随着生物系统的不同而变化, 因此不建议仅以单一的加载速率进行力谱实验.为了更好地了解生物系统的机械性能, 一般需要在较宽范围的加载速率下对该响应进行表征(Krieg et al. 2014, Medalsy & Müller 2013, Herruzo et al. 2014). ...
Transmembrane helices have rough energy surfaces.
1
2007
... 膜蛋白去折叠过程会导致力曲线上产生很多锯齿状的突变峰, 每个突变峰均对应膜蛋白一个片段的去折叠过程.基于Bell-Evans模型推导出了蛋白质去折叠所需力与拉伸速率之间的关系, 揭示了蛋白质力学强度的动力学特性. 当拉伸速率较低时, 去折叠力将正比于拉伸速率, 对于较大的拉伸速率, 去折叠力与拉伸速率成指数关系(Bell 1978, Evans & Ritchie 1997). 通常利用虫链模型(worm-like chain)对力曲线上的各个突变峰进行拟合, 在虫链模型WLC拟合所得到的4个参数中, 每个结构域的能量势垒\(\DeltaG_{0}^\ddag \)的大小反映了跨越该能量势垒的难度, 此值越大越需要更大的作用力去克服, 可以反映结构域的稳定性; 活化态到结合态的距离\(x_{\rm u}\)反映了结构域能够容纳的构象变化, 此值越大则代表在去折叠过程中结构域可能出现的构象形态越多; 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\), 此值越大说明在自然条件下发生解离的速率越大, 结构域不稳定性越高; 等效弹性系数\(\kappa \)反映了结构域的机械力学参数, 此值越大表示结构域的机械硬度越强.通过上述参数可以描绘出去折叠动态过程的能量图谱(Janovjak et al. 2004, Kuo et al. 2010, Schoeler et al. 2015).也有研究表明这一能量图谱的表面粗糙而非平滑(Janovjak et al. 2007).近年来随着模型的优化已经有高度集成化的分析软件(Lamour et al. 2014), 同时随着设备性能的提升, 已经逐渐展开了这一模型在高速拉伸下的分析(Junker & Rief 2010, Berkovich et al. 2012). ...
Probing the energy landscape of the membrane protein bacteriorhodopsin.
1
2004
... 膜蛋白去折叠过程会导致力曲线上产生很多锯齿状的突变峰, 每个突变峰均对应膜蛋白一个片段的去折叠过程.基于Bell-Evans模型推导出了蛋白质去折叠所需力与拉伸速率之间的关系, 揭示了蛋白质力学强度的动力学特性. 当拉伸速率较低时, 去折叠力将正比于拉伸速率, 对于较大的拉伸速率, 去折叠力与拉伸速率成指数关系(Bell 1978, Evans & Ritchie 1997). 通常利用虫链模型(worm-like chain)对力曲线上的各个突变峰进行拟合, 在虫链模型WLC拟合所得到的4个参数中, 每个结构域的能量势垒\(\DeltaG_{0}^\ddag \)的大小反映了跨越该能量势垒的难度, 此值越大越需要更大的作用力去克服, 可以反映结构域的稳定性; 活化态到结合态的距离\(x_{\rm u}\)反映了结构域能够容纳的构象变化, 此值越大则代表在去折叠过程中结构域可能出现的构象形态越多; 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\), 此值越大说明在自然条件下发生解离的速率越大, 结构域不稳定性越高; 等效弹性系数\(\kappa \)反映了结构域的机械力学参数, 此值越大表示结构域的机械硬度越强.通过上述参数可以描绘出去折叠动态过程的能量图谱(Janovjak et al. 2004, Kuo et al. 2010, Schoeler et al. 2015).也有研究表明这一能量图谱的表面粗糙而非平滑(Janovjak et al. 2007).近年来随着模型的优化已经有高度集成化的分析软件(Lamour et al. 2014), 同时随着设备性能的提升, 已经逐渐展开了这一模型在高速拉伸下的分析(Junker & Rief 2010, Berkovich et al. 2012). ...
Force spectroscopy of molecular systems-single molecule spectroscopy of polymers and biomolecules.
... 在细胞力学性质的研究中, 探索细胞杨氏模量随环境参数的变化是非常重要的一类实验.杨氏模量是弹性材料在一定的弹性范围内, 正向应力与正向应变的比值, 它反映材料的刚度. 杨氏模量初期主要应用于物理学领域, 目前在医学和生物学领域也得到了广泛应用.细胞的杨氏模量的各种拟合模型, 主要包括赫兹模型(Johnson et al. 1987)、Sneddon模型(Sneddon et al. 1965)、DMT(Derjaguin-Müller-Toporov)模型(Derjaguin et al. 1975)以及JKR模型(Johnson et al. 1971). ...
Surface energy and the contact of elastic solids.
1
1971
... 在细胞力学性质的研究中, 探索细胞杨氏模量随环境参数的变化是非常重要的一类实验.杨氏模量是弹性材料在一定的弹性范围内, 正向应力与正向应变的比值, 它反映材料的刚度. 杨氏模量初期主要应用于物理学领域, 目前在医学和生物学领域也得到了广泛应用.细胞的杨氏模量的各种拟合模型, 主要包括赫兹模型(Johnson et al. 1987)、Sneddon模型(Sneddon et al. 1965)、DMT(Derjaguin-Müller-Toporov)模型(Derjaguin et al. 1975)以及JKR模型(Johnson et al. 1971). ...
Evidence for a Broad Transition-State Ensemble in Calmodulin Folding from Single-Molecule Force Spectroscopy.
1
2010
... 膜蛋白去折叠过程会导致力曲线上产生很多锯齿状的突变峰, 每个突变峰均对应膜蛋白一个片段的去折叠过程.基于Bell-Evans模型推导出了蛋白质去折叠所需力与拉伸速率之间的关系, 揭示了蛋白质力学强度的动力学特性. 当拉伸速率较低时, 去折叠力将正比于拉伸速率, 对于较大的拉伸速率, 去折叠力与拉伸速率成指数关系(Bell 1978, Evans & Ritchie 1997). 通常利用虫链模型(worm-like chain)对力曲线上的各个突变峰进行拟合, 在虫链模型WLC拟合所得到的4个参数中, 每个结构域的能量势垒\(\DeltaG_{0}^\ddag \)的大小反映了跨越该能量势垒的难度, 此值越大越需要更大的作用力去克服, 可以反映结构域的稳定性; 活化态到结合态的距离\(x_{\rm u}\)反映了结构域能够容纳的构象变化, 此值越大则代表在去折叠过程中结构域可能出现的构象形态越多; 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\), 此值越大说明在自然条件下发生解离的速率越大, 结构域不稳定性越高; 等效弹性系数\(\kappa \)反映了结构域的机械力学参数, 此值越大表示结构域的机械硬度越强.通过上述参数可以描绘出去折叠动态过程的能量图谱(Janovjak et al. 2004, Kuo et al. 2010, Schoeler et al. 2015).也有研究表明这一能量图谱的表面粗糙而非平滑(Janovjak et al. 2007).近年来随着模型的优化已经有高度集成化的分析软件(Lamour et al. 2014), 同时随着设备性能的提升, 已经逐渐展开了这一模型在高速拉伸下的分析(Junker & Rief 2010, Berkovich et al. 2012). ...
Tuning membrane protein mobility by confinement into nanodomains.
1
2016
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
Probing nanomechanical properties from biomolecules to living cells.
Ultrastable atomic force microscopy: Atomic-scale stability and registration in ambient conditions.
1
2009
... 结合力谱技术的原子力显微镜成像模式在扫描时会记录数十万条力曲线数据, 用以反映机械性能或样品间的相互作用.因此缩短单条力曲线数据的记录时间, 能够减少图像获取时间并动态记录生物学的过程.然而有些配体--受体键合或者化学相互作用在非常快的外力作用下, 是远离平衡态的破坏状态, 此时想要推导其平衡态的力学行为是比较困难的(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).因为在高加载速率下, 必须考虑悬臂梁的流体动力学阻力和物理限制(Amo & Garcia 2016). 所以想要反映这类目标接近平衡态时的相互作用, 需要缓慢地施加外力. 因此, 似乎有可能需要开发能够在非常大的范围选择加载速率的方法, 例如结合当前的超稳定AFM方向(King et al. 2009, Bull et al. 2014, Weafer et al. 2012)与超高速AFM方向(Dong et al. 2011, Rico et al. 2013). ...
Video imaging of walking myosin V by high-speed atomic force microscopy.
1
2010
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
Contact mechanics and tip shape in AFM-based nanomechanical measurements.
b. Discriminating small molecule DNA binding modes by single molecule force spectroscopy.
2
2002
... 原子力显微镜力谱技术作用于核酸的各类实验设计. (a)外力诱导下的DNA双螺旋熔融(Krautbauer et al. 2002b), (b)单链DNA与表面的摩擦力测试(Kühner et al. 2006), (c)将RNA链从病毒中直接提取的力学实验(Liu et al. 2010b), (d)外力诱导的DNA双螺旋熔融, 及其在SSB蛋白辅助下的复原(Liu et al. 2010a) ...
... DNA的力曲线具有指纹特征, 只要是双螺旋DNA在拉伸条件下都会存在B-OS 转变平台.而许多DNA结合分子与DNA作用后会引起该转变区域指纹谱的变化, 因此该指纹谱是研究DNA结合试剂(如蛋白质和药物)与DNA相互作用的指示标志(Krautbauer et al.2002b, Krautbauer et al. 2000).原子力显微镜力谱实验中, 探针相对于样品的移动可以不限于在同一点的位置上仅沿\(Z\)轴上下移动, 在系统程序设定的路径下其活动范围可以是整个三维空间. 图6(b)描述了力谱技术用于研究不同聚合物样品(单链DNA, 聚烯丙胺)与云母表面的摩擦力测试(Kühner et al. 2006). 在实验过程中, 探针末端按照指定的路线接近或远离基底表面, 聚合物样品将表现出与表面解离或在表面滑动的现象, 也有可能同时发生这两种现象.于是此时获得的力谱信息, 将能够得到在设定的实验条件下聚合物链样品的摩擦力强弱的信息. 该研究表明, 即便两种聚合物链样品与同一表面具有类似大小的结合力, 它们与表面的摩擦力所反映的面内移动能力的差异仍然有可能具有十分显著的差异. ...
Unzipping DNA oligomers.
1
2003
... 核酸本身的双螺旋结构即可作为配对目标的一种典型研究.通过将单链DNA修饰在原子力显微镜的针尖, 对其与互补DNA单链的相互作用展开了一系列研究(Moy et al. 1995).检测出双链DNA的解链作用力大小为每10碱基对10 \(\sim\) 20 pN(Krautbauer et al. 2003).通过对两个30碱基的寡核苷酸双链体组成的分子力平衡的实验, 给出了一种能够以设定的外力和加载速率模拟力诱导下的断裂模型.并用DNA双链两端双键的断裂位点进行了预测(Neuert et al. 2007).核酸正在越来越多地作为纳米尺度结构的可编程构建块.通过特定DNA碱基对识别进行自组装, 可以形成各种特异性结构.这些结构的稳定化可以通过与取代碱基的金属配位键来实现.在携带水杨醛核苷的双链DNA在铜金属络合时的力学转换研究中, 发现断键力增大了两倍. 动力学分析表明在外力加载速率较高时, 此生物分子杂交体的力学性质主要受局域配位键的影响, 而在外力加载速率较低时, 目标的稳定性主要受碱基对相互结合的影响(Gaub et al. 2009). ...
a. Tensile forces govern germ-layer organization in zebrafish.
0
2008
Mechanical control of the sense of touch by beta-spectrin.
1
2014
... 力谱技术为表征生物系统的机械特性提供了有力的工具, 但是理论推导和实验结果均表明, 这些系统的机械响应是取决于施加力的加载速率(loading rate) (Janmey et al.2007, Hinterdorfer & Dufrêne 2006).加载速率是指施加于被测物体的作用力随时间的变化率, 以某一恒定速率拉回探针时测定的分子间解离力, 只是一个连续的分子间作用力谱中的一个点.由于系统的机械响应与作用力的加载速率是非线性关系, 并且随着生物系统的不同而变化, 因此不建议仅以单一的加载速率进行力谱实验.为了更好地了解生物系统的机械性能, 一般需要在较宽范围的加载速率下对该响应进行表征(Krieg et al. 2014, Medalsy & Müller 2013, Herruzo et al. 2014). ...
b. A bond for a lifetime: employing membrane nanotubes from living cells to determine receptor-ligand kinetics.
1
2008
... 随着研究领域的深入, 人们意识到测量不同分离速度(即加载速率)的断裂力提供了关于结合键的动力学性质的信息(Moy et al.1994).结合键的动力学模型分析可以用于估计配体--受体键的热力学和动力学性质(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).这些性质包括跨越折叠状态的能量势垒\(\Delta G_{0}^\ddag \); 活化态到结合态的距离\(x_{\rm u}\); 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\); 此外结合键的寿命也可以通过使键处于低恒定力并等待键的断裂来测量(Woodside & Block 2014, Perez-Jimenez et al. 2011).此恒定作用力可以通过原子力显微镜的反馈系统以力钳模式施加(Oberhauser et al.2001, Stahl et al. 2009), 也可以利用天然的细胞膜栓系Tether来实现(Krieg et al. 2008b). ...
Friction of single polymers at surfaces.
2
2006
... 原子力显微镜力谱技术作用于核酸的各类实验设计. (a)外力诱导下的DNA双螺旋熔融(Krautbauer et al. 2002b), (b)单链DNA与表面的摩擦力测试(Kühner et al. 2006), (c)将RNA链从病毒中直接提取的力学实验(Liu et al. 2010b), (d)外力诱导的DNA双螺旋熔融, 及其在SSB蛋白辅助下的复原(Liu et al. 2010a) ...
... DNA的力曲线具有指纹特征, 只要是双螺旋DNA在拉伸条件下都会存在B-OS 转变平台.而许多DNA结合分子与DNA作用后会引起该转变区域指纹谱的变化, 因此该指纹谱是研究DNA结合试剂(如蛋白质和药物)与DNA相互作用的指示标志(Krautbauer et al.2002b, Krautbauer et al. 2000).原子力显微镜力谱实验中, 探针相对于样品的移动可以不限于在同一点的位置上仅沿\(Z\)轴上下移动, 在系统程序设定的路径下其活动范围可以是整个三维空间. 图6(b)描述了力谱技术用于研究不同聚合物样品(单链DNA, 聚烯丙胺)与云母表面的摩擦力测试(Kühner et al. 2006). 在实验过程中, 探针末端按照指定的路线接近或远离基底表面, 聚合物样品将表现出与表面解离或在表面滑动的现象, 也有可能同时发生这两种现象.于是此时获得的力谱信息, 将能够得到在设定的实验条件下聚合物链样品的摩擦力强弱的信息. 该研究表明, 即便两种聚合物链样品与同一表面具有类似大小的结合力, 它们与表面的摩擦力所反映的面内移动能力的差异仍然有可能具有十分显著的差异. ...
Optically monitoring the mechanical assembly of single molecules.
1
2009
... 在涉及到DNA的复杂力谱力学操纵实验中, 可以利用互补单链DNA之间会形成DNA双螺旋的特点, 将单个目标分子剪切并黏贴到特定的位置, 与传统方法相比将定位精度显著提高到纳米量级(Heucke et al. 2014).此外, 通过使用表面电位循环技术结合力学操纵, 可以将原子力显微镜尖端的单分子DNA沉积在裸金电极上(Erdmann et al. 2010). 应用抗体和抗原肽标签, 单分子黏贴技术可以处理单一蛋白质.蛋白质--DNA复合物可以排列成复杂的样式, 同时保持蛋白质部分的功能.这种用于蛋白质排列的定向组装的单分子方法, 使得基于蛋白质组分的受控系统构建成为可能(Strackharn et al. 2012).经单分子力谱剪切进行的表面组装, 还可以用纳米级精度实时监测该过程.通过全内反射荧光显微镜和原子力显微镜的组合, 可以观察到纳米尺度图案沉积的单个荧光团(Kufer et al. 2009).虽然图案的光学分辨率远低于显微镜, 但可以通过将结合位点定位, 并检测荧光团的光漂白来识别各个染料. 通过这种方法的组合, 可以随意布置单独的染料标记的生物分子以用于特定功能, 例如偶联的荧光团系统或定制的酶级联, 同时辅以纳米级精度的实时监测. ...
Probing static disorder in Arrhenius kinetics by single-molecule force spectroscopy.
1
2010
... 膜蛋白去折叠过程会导致力曲线上产生很多锯齿状的突变峰, 每个突变峰均对应膜蛋白一个片段的去折叠过程.基于Bell-Evans模型推导出了蛋白质去折叠所需力与拉伸速率之间的关系, 揭示了蛋白质力学强度的动力学特性. 当拉伸速率较低时, 去折叠力将正比于拉伸速率, 对于较大的拉伸速率, 去折叠力与拉伸速率成指数关系(Bell 1978, Evans & Ritchie 1997). 通常利用虫链模型(worm-like chain)对力曲线上的各个突变峰进行拟合, 在虫链模型WLC拟合所得到的4个参数中, 每个结构域的能量势垒\(\DeltaG_{0}^\ddag \)的大小反映了跨越该能量势垒的难度, 此值越大越需要更大的作用力去克服, 可以反映结构域的稳定性; 活化态到结合态的距离\(x_{\rm u}\)反映了结构域能够容纳的构象变化, 此值越大则代表在去折叠过程中结构域可能出现的构象形态越多; 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\), 此值越大说明在自然条件下发生解离的速率越大, 结构域不稳定性越高; 等效弹性系数\(\kappa \)反映了结构域的机械力学参数, 此值越大表示结构域的机械硬度越强.通过上述参数可以描绘出去折叠动态过程的能量图谱(Janovjak et al. 2004, Kuo et al. 2010, Schoeler et al. 2015).也有研究表明这一能量图谱的表面粗糙而非平滑(Janovjak et al. 2007).近年来随着模型的优化已经有高度集成化的分析软件(Lamour et al. 2014), 同时随着设备性能的提升, 已经逐渐展开了这一模型在高速拉伸下的分析(Junker & Rief 2010, Berkovich et al. 2012). ...
Atomic force microscopy in imaging of viruses and virus-infected cells.
2
2011
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
... 原子力显微镜用于各尺寸生物样品成像示例. (a)核酸(Pyne et al. 2014); (b)蛋白质(Seelert et al. 2000); (c)纤维(Stamov et al. 2015); (d)病毒(Kuznetsov & McPherson 2011); (e)细胞(Friedrichs et al. 2007); (f)秀丽线虫横截面, 扫描范围24 \(\mu \)m \(\times \) 24 \(\mu \)m \(\times\) 6um (NT-MDT公司) ...
Atomic force microscopy probing of cell elasticity.
1
2007
... 赫兹模型成立的两个前提条件是压入物呈抛物线形和样品厚度足够大(相对于针尖压细胞的深度), 同时要求样品表面连续且光滑, 实验过程中样品形变量相对较小(Kuznetsova et al. 2007, Costa 2003).当探针为球形时, 赫兹模型为 \[ f_{\rm sphere} =\dfrac{4}{3}\dfrac{E}{1 - v^2}\sqrt R \delta ^{3 / 2} \ \ (7)\] ...
Atomic force microscopy-based single-cell indentation: experimentation and finite element simulation. Proceedings of IEEE// International Conference on Intelligent Robots and Systems,
Easyworm: An open-source software tool to determine the mechanical properties of worm-like chains.
1
2014
... 膜蛋白去折叠过程会导致力曲线上产生很多锯齿状的突变峰, 每个突变峰均对应膜蛋白一个片段的去折叠过程.基于Bell-Evans模型推导出了蛋白质去折叠所需力与拉伸速率之间的关系, 揭示了蛋白质力学强度的动力学特性. 当拉伸速率较低时, 去折叠力将正比于拉伸速率, 对于较大的拉伸速率, 去折叠力与拉伸速率成指数关系(Bell 1978, Evans & Ritchie 1997). 通常利用虫链模型(worm-like chain)对力曲线上的各个突变峰进行拟合, 在虫链模型WLC拟合所得到的4个参数中, 每个结构域的能量势垒\(\DeltaG_{0}^\ddag \)的大小反映了跨越该能量势垒的难度, 此值越大越需要更大的作用力去克服, 可以反映结构域的稳定性; 活化态到结合态的距离\(x_{\rm u}\)反映了结构域能够容纳的构象变化, 此值越大则代表在去折叠过程中结构域可能出现的构象形态越多; 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\), 此值越大说明在自然条件下发生解离的速率越大, 结构域不稳定性越高; 等效弹性系数\(\kappa \)反映了结构域的机械力学参数, 此值越大表示结构域的机械硬度越强.通过上述参数可以描绘出去折叠动态过程的能量图谱(Janovjak et al. 2004, Kuo et al. 2010, Schoeler et al. 2015).也有研究表明这一能量图谱的表面粗糙而非平滑(Janovjak et al. 2007).近年来随着模型的优化已经有高度集成化的分析软件(Lamour et al. 2014), 同时随着设备性能的提升, 已经逐渐展开了这一模型在高速拉伸下的分析(Junker & Rief 2010, Berkovich et al. 2012). ...
Analysis of the effect of LRP-1 silencing on the invasive potential of cancer cells by nanomechanical probing and adhesion force measurements using atomic force microscopy.
Observing the growth of individual actin filaments in cell extracts by time-lapse atomic force microscopy.
1
2003
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Engineering proteins with tailored nanomechanical properties a single molecule approach.
1
2007
... 单分子力谱可以直接探测弹性蛋白质在单分子水平上的机械性能, 并揭示弹性蛋白质分子设计原理. 结合单分子力谱和蛋白质工程技术, 已经可以使用定制的纳米力学性质来设计蛋白质(Li et al. 2007).研究发现, 人工合成的弹性蛋白质, 如GB1蛋白质链, 其力学性质类似甚至超过了天然的蛋白质(Cao & Li 2007).为了利用蛋白质构建人工新型材料, 将其应用到纳米力学系统中作为弹簧、转换器或者感应器, 需要设计一系列具有特定纳米力学性质的人造弹性蛋白质, 例如力学稳定性、延展性和柔韧性等(Lü et al. 2010). 总之, 在纳米尺度下理解它们的结构特性和力学性质, 才能让我们更有效和全面地利用它们.蛋白质力学特性的单分子力谱为此提供了有效途径. ...
Point mutations alter the mechanical stability of immunoglobulin modules.
Water's role in the force-induced unfolding of ubiquitin.
1
2010
... 生物大分子在生理条件下(酸碱度近中性的室温稀盐水溶液)往往会形成规整、精确的超分子结构, 比如双链DNA 以双螺旋的形式存在, 蛋白质则经由\(\alpha \)-螺旋、\(\beta\)-折叠等结构完成组装. 为了研究生物大分子与水的相互作用, Cui等利用单分子力谱迫使高分子单链进入到不良溶剂中, 开展了生物大分子在水溶液与非极性溶剂中的对照研究(Cui et al. 2006, 2007). 双链DNA 在受到外力拉伸时, 可在约65 pN处观察到一个长平台, 对应于从B型双链DNA到其过度拉伸态的结构转变. 然而实验中发现, 在不同溶剂中力曲线存在着明显的变化.如果将水溶液替换为非极性溶剂(如二乙苯等), 不仅无法观测到指纹特征, 而且所得力曲线与单链DNA的一致.这一结果表明双链DNA在非极性溶剂中将变性为单链结构.在实验结果的基础上, 提出ssDNA与水分子之间的弱氢键可能是双链DNA稳定存在于水中的前提条件.原子力显微镜也被广泛应用与除DNA之外的许多小分子的力谱研究, 甚至包括了金颗粒(Xue et al. 2014)、纳米线(Marszalek et al. 2000)等非生物样品, 如图7(b)所示.早期的研究应用实例包括应用力谱分析技术在单分子水平上鉴定多糖混合物的组分(Marszalek et al.2001), 观察黄原胶分子与基底的结合力(Li et al. 1999a), 研究蚕丝素分子的纳米力学特性(Zhang et al. 2000), 观察肝素分子在外力作用下的力学性质(Marszalek et al. 2003), Fern\'{a}ndez课题组对泛素分子力学性质、及其与水分子相互作用的一系列研究等(Carrion-Vazquez et al.2003, Schlierf & Fern\'{a}ndez 2004, Brujić et al. 2006, Li et al. 2010). ...
Molecular recognition force spectroscopy study of the dynamic interaction between aptamer GBI-10 and extracellular matrix protein tenascin-C on human glioblastoma cell.
Kinetic measurements on single-molecule disulfide bond cleavage.
0
2011
Investigation of free fatty acid associated recombinant membrane receptor protein expression in HEK293 cells using raman spectroscopy, calcium imaging, and atomic force microscopy.
Extracting a single polyethylene oxide chain from a single crystal by a combination of atomic force microscopy imaging and single-molecule force spectroscopy: toward the investigation of molecular interactions in their condensed states.
1
2011
... 原子力显微镜力谱技术作用于小分子的各类实验设计. (a)低聚乙二醇共聚物长链分子在不用缓冲液成分下与金膜的结合力分析实验(Nash & Gaub 2012), (b)金颗粒纳米棒在探针作用下的力学及导电性分析实验(Marszalek et al. 2000), (c) 聚氧乙烯(PEO)链在其单晶中的折叠样式分析实验(Liu et al. 2011), (d)光敏分子受光照激发产生形变所带动的力谱分析实验(Hugel et al. 2002) ...
a. The nature of the force-induced conformation transition of dsDNA studied by using single molecule force spectroscopy.
2
2010
... 原子力显微镜力谱技术作用于核酸的各类实验设计. (a)外力诱导下的DNA双螺旋熔融(Krautbauer et al. 2002b), (b)单链DNA与表面的摩擦力测试(Kühner et al. 2006), (c)将RNA链从病毒中直接提取的力学实验(Liu et al. 2010b), (d)外力诱导的DNA双螺旋熔融, 及其在SSB蛋白辅助下的复原(Liu et al. 2010a) ...
b. Pulling genetic RNA out of tobacco mosaic virus using single-molecule force spectroscopy.
2
2010
... 原子力显微镜力谱技术作用于核酸的各类实验设计. (a)外力诱导下的DNA双螺旋熔融(Krautbauer et al. 2002b), (b)单链DNA与表面的摩擦力测试(Kühner et al. 2006), (c)将RNA链从病毒中直接提取的力学实验(Liu et al. 2010b), (d)外力诱导的DNA双螺旋熔融, 及其在SSB蛋白辅助下的复原(Liu et al. 2010a) ...
Planned nanostructures of colloidal gold via self-assembly on hierarchically assembled organic bilayer template patterns with in-situ generated terminal amino functionality.
1
2004
... 除此以外, 通过基于力谱技术的原子力显微镜设备, 可以精确控制悬臂梁和探针在样品上移动轨迹和所施加的作用力.再加上可以选择具有不同物理性质的探针, 如镀有铂、金、钛等导电层或者镀有类金刚石耐磨层, 辅以施加在探针或样品上的电压, 还可以进行刻蚀和操纵实验等(Pires et al. 2010). 既可以应用于如图14所示的金属或半导体基底, 也可以用于生物样品表面的刻蚀和特定结构刻画(Liu et al. 2004, Paul et al.2011). 对基于原子力显微镜的力学操纵和刻蚀技术, Garcia等(2014)的综述文献中进行了详细报道. ...
Designed biomaterials to mimic the mechanical properties of muscles.
1
2010
... 单分子力谱可以直接探测弹性蛋白质在单分子水平上的机械性能, 并揭示弹性蛋白质分子设计原理. 结合单分子力谱和蛋白质工程技术, 已经可以使用定制的纳米力学性质来设计蛋白质(Li et al. 2007).研究发现, 人工合成的弹性蛋白质, 如GB1蛋白质链, 其力学性质类似甚至超过了天然的蛋白质(Cao & Li 2007).为了利用蛋白质构建人工新型材料, 将其应用到纳米力学系统中作为弹簧、转换器或者感应器, 需要设计一系列具有特定纳米力学性质的人造弹性蛋白质, 例如力学稳定性、延展性和柔韧性等(Lü et al. 2010). 总之, 在纳米尺度下理解它们的结构特性和力学性质, 才能让我们更有效和全面地利用它们.蛋白质力学特性的单分子力谱为此提供了有效途径. ...
Design and facile production of recombinant resilin-like polypeptides: Gene construction and a rapid protein purification method.
1
2007
... 弹性蛋白质在生物力学系统中是一个重要的功能基本单元, 因为它们具有理想的弹性力学强度和抗性. 弹性蛋白质是天然黏合剂, 可以作为细胞黏附和肌肉蛋白质弹性的基础, 也可以作为具有极好机械性能的结构材料.随着对蛋白质力学性质了解的不断深入, 采用蛋白质人工构建新型材料的研究有了重要的进展(Guerette et al. 1996, Smith et al. 1999, Ardell & Andersen 2001, Becker et al. 2003, Elvin et al. 2005, Lyons et al. 2007).例如人们已经能够利用已知的蛛丝蛋白质的结构合成具有非常好的延展性和强度的人工材料(Wong et al.2012), 这种方法也为制造下一代力学稳定的生物材料指明了方向(Heim et al. 2009). ...
Visualization of DNA and protein-DNA complexes with atomic force microscopy.
Atomic force microscopy observation and characterization of single virions and virus-like particles by nano-indentation.
1
2016
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
pH{\-}induced conformational change of the beta-barrel forming protein OmpG reconstituted into native E. coli lipids.
1
2010
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Gating of the MlotiK1 potassium channel involves large rearrangements of the cyclic nucleotide-binding domains.
2
2011
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
... )、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Atomic force microscopy captures quantized plastic deformation in gold nanowires.
2
2000
... 生物大分子在生理条件下(酸碱度近中性的室温稀盐水溶液)往往会形成规整、精确的超分子结构, 比如双链DNA 以双螺旋的形式存在, 蛋白质则经由\(\alpha \)-螺旋、\(\beta\)-折叠等结构完成组装. 为了研究生物大分子与水的相互作用, Cui等利用单分子力谱迫使高分子单链进入到不良溶剂中, 开展了生物大分子在水溶液与非极性溶剂中的对照研究(Cui et al. 2006, 2007). 双链DNA 在受到外力拉伸时, 可在约65 pN处观察到一个长平台, 对应于从B型双链DNA到其过度拉伸态的结构转变. 然而实验中发现, 在不同溶剂中力曲线存在着明显的变化.如果将水溶液替换为非极性溶剂(如二乙苯等), 不仅无法观测到指纹特征, 而且所得力曲线与单链DNA的一致.这一结果表明双链DNA在非极性溶剂中将变性为单链结构.在实验结果的基础上, 提出ssDNA与水分子之间的弱氢键可能是双链DNA稳定存在于水中的前提条件.原子力显微镜也被广泛应用与除DNA之外的许多小分子的力谱研究, 甚至包括了金颗粒(Xue et al. 2014)、纳米线(Marszalek et al. 2000)等非生物样品, 如图7(b)所示.早期的研究应用实例包括应用力谱分析技术在单分子水平上鉴定多糖混合物的组分(Marszalek et al.2001), 观察黄原胶分子与基底的结合力(Li et al. 1999a), 研究蚕丝素分子的纳米力学特性(Zhang et al. 2000), 观察肝素分子在外力作用下的力学性质(Marszalek et al. 2003), Fern\'{a}ndez课题组对泛素分子力学性质、及其与水分子相互作用的一系列研究等(Carrion-Vazquez et al.2003, Schlierf & Fern\'{a}ndez 2004, Brujić et al. 2006, Li et al. 2010). ...
... 原子力显微镜力谱技术作用于小分子的各类实验设计. (a)低聚乙二醇共聚物长链分子在不用缓冲液成分下与金膜的结合力分析实验(Nash & Gaub 2012), (b)金颗粒纳米棒在探针作用下的力学及导电性分析实验(Marszalek et al. 2000), (c) 聚氧乙烯(PEO)链在其单晶中的折叠样式分析实验(Liu et al. 2011), (d)光敏分子受光照激发产生形变所带动的力谱分析实验(Hugel et al. 2002) ...
Mechanical unfolding intermediates in titin modules.
1
1999
... 此后单个蛋白质去折叠过程的研究蓬勃发展, 对肌联蛋白展开的后续研究发现了其去折叠过程的中间态, 并描绘了其动力学能谱(Marszalek et al. 1999, Schlierf & Rief 2006), Li 等(2002)在此基础上将Titin 蛋白质分子作为一个弹簧处理, 从总体上了解其机械特性.笔者的实验进一步证明了去折叠过程中间态由于其与外力加载速率高度独立的性质, 起到了"力学缓冲"的作用, 其建立的动力学能量壁垒大大延长了I27蛋白质的寿命.实验数据建立了肌联蛋白动力学和拓扑结构间的联系.诸如I27的肌联蛋白末端结构域能够缓冲外力, 分子动力学模拟显示连接\(\beta \)折叠A的氢键断裂时形成了去折叠中间态.末端Ig蛋白质中, 连接\(\beta \)折叠A, B, G的氢键相互作用力, 能够在生理环境所能施加的外力加载速率下, 维持Ig蛋白质折叠结构能垒的稳定. 因此, 能够使蛋白质进入去折叠过程的作用力保持恒定.这一末端Ig中的机制能够使结构域在生理环境产生的外力下保持稳定, 从而在宏观上使肌联蛋白具有了外力吸收能力, 显著降低了意外进入去折叠过程的几率(Nunes et al. 2010).随着设备的进步, 目前已经可以在接近分子动态模拟的时间尺度上进行快速的去折叠实验(Rico et al. 2013). ...
Polysaccharide elasticity governed by chair-boat transitions of the glucopyranose ring.
Direct high-resolution measurement of furrow stiffening during division of adherent cells.
1
2001
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
Nanomechanical properties of proteins and membranes depend on loading rate and electrostatic interactions.
3
2013
... 力谱技术为表征生物系统的机械特性提供了有力的工具, 但是理论推导和实验结果均表明, 这些系统的机械响应是取决于施加力的加载速率(loading rate) (Janmey et al.2007, Hinterdorfer & Dufrêne 2006).加载速率是指施加于被测物体的作用力随时间的变化率, 以某一恒定速率拉回探针时测定的分子间解离力, 只是一个连续的分子间作用力谱中的一个点.由于系统的机械响应与作用力的加载速率是非线性关系, 并且随着生物系统的不同而变化, 因此不建议仅以单一的加载速率进行力谱实验.为了更好地了解生物系统的机械性能, 一般需要在较宽范围的加载速率下对该响应进行表征(Krieg et al. 2014, Medalsy & Müller 2013, Herruzo et al. 2014). ...
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
... 这些早期的实验主要受到设备采样率和压电陶瓷性能的限制, 其每条力曲线都需要接近数秒钟的时间完成, 这就大大制约了能够采集的力曲线的数量.由于每个像素至少要有一条力曲线才能计算机械性能, 所以早期的功能探针力谱结合成像技术结果往往像素点都比较少, 且实验时间比较长. 最近五年来, 随着设备性能的提升、新模式的推广, 以及探针相对样品运动轨迹的改良, 商品化设备的力曲线采集频率已经可以提升到2 kHz左右, 使得配对样品的高通量表征成为了现实(见图21).这方面的应用实例包括: 生物膜和膜蛋白的静电性质(Medalsy & Müller 2013, Guo et al. 2016, Pfreundschuh et al. 2013), 被细菌排出的噬菌体(Alsteens et al. 2013a), 结合动物细胞表面受体的包膜病毒(Alsteens et al. 2017b), 结合人G蛋白偶联受体的配体(Alsteens et al. 2015), 两个配体与相同受体的结合(Pfreundschuh et al. 2015), 以及大型蛋白质复合物上配体结合事件的高清成像等(Pfreundschuh et al. 2014, Kim et al. 2015). ...
Imaging and quantifying chemical and physical properties of native proteins at molecular resolution by force-volume AFM.
1
2011
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy.
1
1999
... 测量断键力随外力加载速率的变化, 便可获得动力学力谱(dynamic force spectroscopy), 并计算出在不施加外力的平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\)、平均寿命\(t_{0}\)、活化态到结合态的距离\(x_{\rm u}\)等动力学参数, 以及复合物在解离过程中要跨越几个势垒, 是否存在复合物解离的中间体等(Lee et al. 2007, Strunz et al. 2000).单分子力谱是单分子水平研究生物分子复合物解离途径和解离动力学过程的有效方法.根据Bell-Evans模型, 施加复合物的断键方向上的力会减小复合物解离的活化能, 增大其解离速率常数, 单对分子复合物的断键力\(F\)与ln\(r\)\((r\)为外力加载速率)之间存在线性关系(Bell 1978, Evans & Ritchie 1997). 此外, 单分子力谱也可用来估计针尖上的配体与基底上受体之间的结合速率常数\(k_{\rm on}\)(Baumgartner et al. 2000).测量不同作用力下单分子相互作用的寿命, 就可获得势垒的位置与高度, 结合平衡态的热力学研究, 就可较完整地描绘出能量形貌图(Merkel et al. 1999). ...
Revealing the bifurcation in the unfolding pathways of GFP by using single-molecule experiments and simulations.
1
2007
... 对于较大的蛋白质, 其经常表现出多个承受力单元, 使得蛋白质的去折叠不是"全或无"的过程, 而是显示出一系列的中间态, 比如绿色荧光蛋白(GFP) (Bertz et al. 2008, Mickler et al. 2007)、强化黄色荧光蛋白(EYFP) (Perez-Jimenez et al. 2006)、T4溶菌酶(T4 lysozyme) (Peng & Li 2008)、艾滋病毒1型受体蛋白CD4(Perez-Jimenez et al. 2014)、人体晶状蛋白(Garcia-Manyes et al. 2015)、麦芽糖结合蛋白(maltose binding protein) (Aggarwal et al. 2011)以及巨肌蛋白激酶(titin kinase)等(Puchner & Gaub 2009, Pernigo et al. 2010). ...
In vivo analysis of local wall stiffness at the shoot apical meristem in Arabidopsis using atomic force microscopy.
Voltage and pH{\-}induced channel closure of porin OmpF visualized by atomic force microscopy.
1
1999
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Conformational changes in surface structures of isolated Connexin26 gap junctions.
1
2002
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Single-molecule adhesion of a stimuli-responsive oligo(ethylene glycol) copolymer to gold.
2
2012
... 原子力显微镜力谱技术作用于小分子的各类实验设计. (a)低聚乙二醇共聚物长链分子在不用缓冲液成分下与金膜的结合力分析实验(Nash & Gaub 2012), (b)金颗粒纳米棒在探针作用下的力学及导电性分析实验(Marszalek et al. 2000), (c) 聚氧乙烯(PEO)链在其单晶中的折叠样式分析实验(Liu et al. 2011), (d)光敏分子受光照激发产生形变所带动的力谱分析实验(Hugel et al. 2002) ...
... 此外, Sonnenberg等对聚丙烯酸单层进行了力谱实验, 观察其相对缓冲液pH值的函数, 以研究连接着基底与针尖两个表面的聚合物的黏附性质(Sonnenberg et al. 2007a). 该研究表明, 聚丙烯酸链和氮化硅探针针尖之间的相互作用力取决于聚合物单层的覆盖密度, 以及聚合物链的轮廓长度.聚合物桥连的稳定性由两个表面的黏合强度决定, 作为依赖性聚电解质单层, 这一强度可以通过使用缓冲液pH值来进行调节.Zhang课题组使用了单分子力谱技术结合受控分子动力学(steered molecular dynamics, SMD)模拟的组合, 研究聚氧乙烯(PEO)单晶中高分子链之间的相互作用, 以及聚氧乙烯(PEO)链在其单晶中的折叠样式, 如图7(c)所示(Song et al.2013, Liu et al. 2011). 研究结果表明, 将单个PEO链从其单晶中提拉出来的作用力大小约为40 pN, 在稀溶液中形成的晶体中的PEO链的折叠样式遵循就近排列的重入折叠模型.在从熔体获得的晶体中, 力曲线上出现的大幅度波动主要是由于折叠样式上大而无规律的环状结构引起的.Liang和Fern\'{a}ndez(2011)的工作研究了单分子亲核取代化学反应中机械能与热效应之间的关系.通过单分子力谱技术研究了TCEP, 1, 4-\(_{\rm DL}\)-DTT和氢硫化物阴离子, 对受机械拉伸作用的多肽二硫键的影响.在一系列精确控制的温度下记录双分子亲核取代(S\(_{\rm N}\)2)事件, 对力和温度引起的反应速率的影响的比较表明, 每增大50 pN的拉力等效于温度升高10 K. Nash和Gaub(2012)测量了单一共聚物分子与金表面的黏附特性, 如图7(a)所示.研究者发现共聚物在高离子浓度缓冲液中的脱水和坍缩, 导致保持分子桥连长度的分布显著降低.在高盐浓度下聚合物显示出较低的绝对解离力, 而协同作用增加了多链相互作用中每条链对应的解离力. 研究结果证实, 坍缩构象中的聚合物在金表面上占据的体积显著减少.这些结果证明在单分子水平下, 溶剂诱导的环境响应共聚物的坍缩能够以可控的方式调节表面黏附力和桥接长度的分布. ...
Predicting the rupture probabilities of molecular bonds in series.
1
2007
... 核酸本身的双螺旋结构即可作为配对目标的一种典型研究.通过将单链DNA修饰在原子力显微镜的针尖, 对其与互补DNA单链的相互作用展开了一系列研究(Moy et al. 1995).检测出双链DNA的解链作用力大小为每10碱基对10 \(\sim\) 20 pN(Krautbauer et al. 2003).通过对两个30碱基的寡核苷酸双链体组成的分子力平衡的实验, 给出了一种能够以设定的外力和加载速率模拟力诱导下的断裂模型.并用DNA双链两端双键的断裂位点进行了预测(Neuert et al. 2007).核酸正在越来越多地作为纳米尺度结构的可编程构建块.通过特定DNA碱基对识别进行自组装, 可以形成各种特异性结构.这些结构的稳定化可以通过与取代碱基的金属配位键来实现.在携带水杨醛核苷的双链DNA在铜金属络合时的力学转换研究中, 发现断键力增大了两倍. 动力学分析表明在外力加载速率较高时, 此生物分子杂交体的力学性质主要受局域配位键的影响, 而在外力加载速率较低时, 目标的稳定性主要受碱基对相互结合的影响(Gaub et al. 2009). ...
Designing an extracellular matrix protein with enhanced mechanical stability.
1
2007
... GB1是一种具有显著机械稳定性的小蛋白质, 其剪切拓扑在确定其机械稳定性方面起着重要的作用.结合单分子力谱和蛋白质工程技术, 研究了侧链还原和疏水核心外壳对GB1机械稳定性的影响.通过设计七点突变体, 对GB1的机械展开进行了分析.研究者发现在疏水核心(F30L/Y45L/F52L)中, 通过施加的拉伸力, 两个剪切的功能域表面上的三个突变导致GB1的机械展开力的显着降低.与野生型GB1相比, 这些突变体的机械展开力下降了50 \(\sim \) 90 pN, 以400 nm/s的拉伸速度时在约180 pN下去折叠. 结果表明, 疏水核心外壳在确定GB1的机械稳定性方面起重要作用, 并表明优化与剪切结构表面的疏水相互作用, 可能是提高GB1和GB1同系物机械稳定性的有效方法(Bu et al. 2012).此外, 在蛋白质受力区域之外的突变也可以降低蛋白质的力学稳定性(Balamurali et al.2008, Ng et al. 2007). ...
A "force buffer" protecting immunoglobulin titin.
1
2010
... 此后单个蛋白质去折叠过程的研究蓬勃发展, 对肌联蛋白展开的后续研究发现了其去折叠过程的中间态, 并描绘了其动力学能谱(Marszalek et al. 1999, Schlierf & Rief 2006), Li 等(2002)在此基础上将Titin 蛋白质分子作为一个弹簧处理, 从总体上了解其机械特性.笔者的实验进一步证明了去折叠过程中间态由于其与外力加载速率高度独立的性质, 起到了"力学缓冲"的作用, 其建立的动力学能量壁垒大大延长了I27蛋白质的寿命.实验数据建立了肌联蛋白动力学和拓扑结构间的联系.诸如I27的肌联蛋白末端结构域能够缓冲外力, 分子动力学模拟显示连接\(\beta \)折叠A的氢键断裂时形成了去折叠中间态.末端Ig蛋白质中, 连接\(\beta \)折叠A, B, G的氢键相互作用力, 能够在生理环境所能施加的外力加载速率下, 维持Ig蛋白质折叠结构能垒的稳定. 因此, 能够使蛋白质进入去折叠过程的作用力保持恒定.这一末端Ig中的机制能够使结构域在生理环境产生的外力下保持稳定, 从而在宏观上使肌联蛋白具有了外力吸收能力, 显著降低了意外进入去折叠过程的几率(Nunes et al. 2010).随着设备的进步, 目前已经可以在接近分子动态模拟的时间尺度上进行快速的去折叠实验(Rico et al. 2013). ...
Action of the Hsp70 chaperone system observed with single proteins.
1
2015
... 总之, 蛋白质选择特异的去折叠途径与其局部电荷、环境以及氨基酸复杂的相互作用有关, 如分子伴侣(Mashaghi et al. 2013, Nunes et al. 2015)或其他辅因子的外部试剂(Puchner & Gaub 2012, Perez-Jimenez et al.2011), 也会对蛋白质的稳定性和折叠途径产生影响. ...
Stepwise unfolding of titin under force-clamp atomic force microscopy.
1
2001
... 随着研究领域的深入, 人们意识到测量不同分离速度(即加载速率)的断裂力提供了关于结合键的动力学性质的信息(Moy et al.1994).结合键的动力学模型分析可以用于估计配体--受体键的热力学和动力学性质(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).这些性质包括跨越折叠状态的能量势垒\(\Delta G_{0}^\ddag \); 活化态到结合态的距离\(x_{\rm u}\); 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\); 此外结合键的寿命也可以通过使键处于低恒定力并等待键的断裂来测量(Woodside & Block 2014, Perez-Jimenez et al. 2011).此恒定作用力可以通过原子力显微镜的反馈系统以力钳模式施加(Oberhauser et al.2001, Stahl et al. 2009), 也可以利用天然的细胞膜栓系Tether来实现(Krieg et al. 2008b). ...
Single protein misfolding events captured by atomic force microscopy.
Detection of TRKB receptors distributed in cultured hippocampal neurons through bioconjugation between highly luminescent (quantum dot-neutravidin) and (biotinylated anti-TRKB antibody) on neurons by combined atomic force microscope and confocal laser scanning microscope.
0
2010
Stabilizing effect of Zn\(^{2 + }\) in native bovine rhodopsin.
0
2007
Rapid turnaround scanning probe nanolithography.
1
2011
... 除此以外, 通过基于力谱技术的原子力显微镜设备, 可以精确控制悬臂梁和探针在样品上移动轨迹和所施加的作用力.再加上可以选择具有不同物理性质的探针, 如镀有铂、金、钛等导电层或者镀有类金刚石耐磨层, 辅以施加在探针或样品上的电压, 还可以进行刻蚀和操纵实验等(Pires et al. 2010). 既可以应用于如图14所示的金属或半导体基底, 也可以用于生物样品表面的刻蚀和特定结构刻画(Liu et al. 2004, Paul et al.2011). 对基于原子力显微镜的力学操纵和刻蚀技术, Garcia等(2014)的综述文献中进行了详细报道. ...
Atomic force microscopy reveals parallel mechanical unfolding pathways of T4 lysozyme: evidence for a kinetic partitioning mechanism.
2
2008
... 对于较大的蛋白质, 其经常表现出多个承受力单元, 使得蛋白质的去折叠不是"全或无"的过程, 而是显示出一系列的中间态, 比如绿色荧光蛋白(GFP) (Bertz et al. 2008, Mickler et al. 2007)、强化黄色荧光蛋白(EYFP) (Perez-Jimenez et al. 2006)、T4溶菌酶(T4 lysozyme) (Peng & Li 2008)、艾滋病毒1型受体蛋白CD4(Perez-Jimenez et al. 2014)、人体晶状蛋白(Garcia-Manyes et al. 2015)、麦芽糖结合蛋白(maltose binding protein) (Aggarwal et al. 2011)以及巨肌蛋白激酶(titin kinase)等(Puchner & Gaub 2009, Pernigo et al. 2010). ...
... 图10所示的实验通过设计蛋白质分子的组成结构, 研究了其对蛋白质力学性质的影响(Cao et al. 2007)单一的NuG2蛋白质串每个结构域去折叠作用力峰值约为100nN, 结合了hFc的NuG2蛋白质串每个结构域去折叠作用力的峰值约为200nN.通过蛋白质工程技术, 以8个NuG2蛋白质串为基底, 在其上选择性地结合上4个hFc蛋白质. 在新的力谱实验中可以发现, 不论哪个NuG2蛋白质单元结合了hFc蛋白质, 力谱曲线中总是会首先出现4个100nN的锯齿峰, 这说明在蛋白质串中总是没有结合hFc蛋白质的单一NuG2蛋白质单元最先去折叠.类似的现象也在T4溶菌酶和GB1蛋白质串的设计中得到验证, 夹在GB1蛋白质串中的T4溶菌酶由于具有更低的结构稳定性, 力谱实验中其对应的锯齿峰也会先于稳定性更高的GB1蛋白质锯齿峰出现(Peng & Li 2008). 此外, 类似的实验设计还包括用GB1蛋白质串串联其他的目标分子, 如GL5/I27w34f; 用GL5蛋白质串串联T4L目标蛋白质; 用Ig27蛋白质串联GV54P目标蛋白质等(Peng & Li 2009a, 2009b; Cao et al. 2008). ...
a. Direct Observation of tug-of-war during the folding of a mutually exclusive protein.
1
2009
... 图10所示的实验通过设计蛋白质分子的组成结构, 研究了其对蛋白质力学性质的影响(Cao et al. 2007)单一的NuG2蛋白质串每个结构域去折叠作用力峰值约为100nN, 结合了hFc的NuG2蛋白质串每个结构域去折叠作用力的峰值约为200nN.通过蛋白质工程技术, 以8个NuG2蛋白质串为基底, 在其上选择性地结合上4个hFc蛋白质. 在新的力谱实验中可以发现, 不论哪个NuG2蛋白质单元结合了hFc蛋白质, 力谱曲线中总是会首先出现4个100nN的锯齿峰, 这说明在蛋白质串中总是没有结合hFc蛋白质的单一NuG2蛋白质单元最先去折叠.类似的现象也在T4溶菌酶和GB1蛋白质串的设计中得到验证, 夹在GB1蛋白质串中的T4溶菌酶由于具有更低的结构稳定性, 力谱实验中其对应的锯齿峰也会先于稳定性更高的GB1蛋白质锯齿峰出现(Peng & Li 2008). 此外, 类似的实验设计还包括用GB1蛋白质串串联其他的目标分子, 如GL5/I27w34f; 用GL5蛋白质串串联T4L目标蛋白质; 用Ig27蛋白质串联GV54P目标蛋白质等(Peng & Li 2009a, 2009b; Cao et al. 2008). ...
b. Domain insertion effectively regulates the mechanical unfolding hierarchy of elastomeric proteins toward engineering multifunctional elastomeric proteins.
1
2009
... 图10所示的实验通过设计蛋白质分子的组成结构, 研究了其对蛋白质力学性质的影响(Cao et al. 2007)单一的NuG2蛋白质串每个结构域去折叠作用力峰值约为100nN, 结合了hFc的NuG2蛋白质串每个结构域去折叠作用力的峰值约为200nN.通过蛋白质工程技术, 以8个NuG2蛋白质串为基底, 在其上选择性地结合上4个hFc蛋白质. 在新的力谱实验中可以发现, 不论哪个NuG2蛋白质单元结合了hFc蛋白质, 力谱曲线中总是会首先出现4个100nN的锯齿峰, 这说明在蛋白质串中总是没有结合hFc蛋白质的单一NuG2蛋白质单元最先去折叠.类似的现象也在T4溶菌酶和GB1蛋白质串的设计中得到验证, 夹在GB1蛋白质串中的T4溶菌酶由于具有更低的结构稳定性, 力谱实验中其对应的锯齿峰也会先于稳定性更高的GB1蛋白质锯齿峰出现(Peng & Li 2008). 此外, 类似的实验设计还包括用GB1蛋白质串串联其他的目标分子, 如GL5/I27w34f; 用GL5蛋白质串串联T4L目标蛋白质; 用Ig27蛋白质串联GV54P目标蛋白质等(Peng & Li 2009a, 2009b; Cao et al. 2008). ...
Probing the effect of force on HIV{\-}1 receptor CD4.
1
2014
... 对于较大的蛋白质, 其经常表现出多个承受力单元, 使得蛋白质的去折叠不是"全或无"的过程, 而是显示出一系列的中间态, 比如绿色荧光蛋白(GFP) (Bertz et al. 2008, Mickler et al. 2007)、强化黄色荧光蛋白(EYFP) (Perez-Jimenez et al. 2006)、T4溶菌酶(T4 lysozyme) (Peng & Li 2008)、艾滋病毒1型受体蛋白CD4(Perez-Jimenez et al. 2014)、人体晶状蛋白(Garcia-Manyes et al. 2015)、麦芽糖结合蛋白(maltose binding protein) (Aggarwal et al. 2011)以及巨肌蛋白激酶(titin kinase)等(Puchner & Gaub 2009, Pernigo et al. 2010). ...
Mechanical unfolding pathways of the enhanced yellow fluorescent protein revealed by single molecule force spectroscopy.
1
2006
... 对于较大的蛋白质, 其经常表现出多个承受力单元, 使得蛋白质的去折叠不是"全或无"的过程, 而是显示出一系列的中间态, 比如绿色荧光蛋白(GFP) (Bertz et al. 2008, Mickler et al. 2007)、强化黄色荧光蛋白(EYFP) (Perez-Jimenez et al. 2006)、T4溶菌酶(T4 lysozyme) (Peng & Li 2008)、艾滋病毒1型受体蛋白CD4(Perez-Jimenez et al. 2014)、人体晶状蛋白(Garcia-Manyes et al. 2015)、麦芽糖结合蛋白(maltose binding protein) (Aggarwal et al. 2011)以及巨肌蛋白激酶(titin kinase)等(Puchner & Gaub 2009, Pernigo et al. 2010). ...
Single-molecule paleoenzymology probes the chemistry of resurrected enzymes.
2
2011
... 总之, 蛋白质选择特异的去折叠途径与其局部电荷、环境以及氨基酸复杂的相互作用有关, 如分子伴侣(Mashaghi et al. 2013, Nunes et al. 2015)或其他辅因子的外部试剂(Puchner & Gaub 2012, Perez-Jimenez et al.2011), 也会对蛋白质的稳定性和折叠途径产生影响. ...
... 随着研究领域的深入, 人们意识到测量不同分离速度(即加载速率)的断裂力提供了关于结合键的动力学性质的信息(Moy et al.1994).结合键的动力学模型分析可以用于估计配体--受体键的热力学和动力学性质(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).这些性质包括跨越折叠状态的能量势垒\(\Delta G_{0}^\ddag \); 活化态到结合态的距离\(x_{\rm u}\); 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\); 此外结合键的寿命也可以通过使键处于低恒定力并等待键的断裂来测量(Woodside & Block 2014, Perez-Jimenez et al. 2011).此恒定作用力可以通过原子力显微镜的反馈系统以力钳模式施加(Oberhauser et al.2001, Stahl et al. 2009), 也可以利用天然的细胞膜栓系Tether来实现(Krieg et al. 2008b). ...
Diversity of chemical mechanisms in thioredoxin catalysis revealed by single-molecule force spectroscopy.
Structural insight into M-band assembly and mechanics from the titin-obscurin-like-1 complex.
1
2010
... 对于较大的蛋白质, 其经常表现出多个承受力单元, 使得蛋白质的去折叠不是"全或无"的过程, 而是显示出一系列的中间态, 比如绿色荧光蛋白(GFP) (Bertz et al. 2008, Mickler et al. 2007)、强化黄色荧光蛋白(EYFP) (Perez-Jimenez et al. 2006)、T4溶菌酶(T4 lysozyme) (Peng & Li 2008)、艾滋病毒1型受体蛋白CD4(Perez-Jimenez et al. 2014)、人体晶状蛋白(Garcia-Manyes et al. 2015)、麦芽糖结合蛋白(maltose binding protein) (Aggarwal et al. 2011)以及巨肌蛋白激酶(titin kinase)等(Puchner & Gaub 2009, Pernigo et al. 2010). ...
Localizing chemical groups while imaging single native proteins by high-resolution atomic force microscopy.
1
2014
... 这些早期的实验主要受到设备采样率和压电陶瓷性能的限制, 其每条力曲线都需要接近数秒钟的时间完成, 这就大大制约了能够采集的力曲线的数量.由于每个像素至少要有一条力曲线才能计算机械性能, 所以早期的功能探针力谱结合成像技术结果往往像素点都比较少, 且实验时间比较长. 最近五年来, 随着设备性能的提升、新模式的推广, 以及探针相对样品运动轨迹的改良, 商品化设备的力曲线采集频率已经可以提升到2 kHz左右, 使得配对样品的高通量表征成为了现实(见图21).这方面的应用实例包括: 生物膜和膜蛋白的静电性质(Medalsy & Müller 2013, Guo et al. 2016, Pfreundschuh et al. 2013), 被细菌排出的噬菌体(Alsteens et al. 2013a), 结合动物细胞表面受体的包膜病毒(Alsteens et al. 2017b), 结合人G蛋白偶联受体的配体(Alsteens et al. 2015), 两个配体与相同受体的结合(Pfreundschuh et al. 2015), 以及大型蛋白质复合物上配体结合事件的高清成像等(Pfreundschuh et al. 2014, Kim et al. 2015). ...
Identifying and quantifying two ligand-binding sites while imaging native human membrane receptors by AFM.
1
2015
... 这些早期的实验主要受到设备采样率和压电陶瓷性能的限制, 其每条力曲线都需要接近数秒钟的时间完成, 这就大大制约了能够采集的力曲线的数量.由于每个像素至少要有一条力曲线才能计算机械性能, 所以早期的功能探针力谱结合成像技术结果往往像素点都比较少, 且实验时间比较长. 最近五年来, 随着设备性能的提升、新模式的推广, 以及探针相对样品运动轨迹的改良, 商品化设备的力曲线采集频率已经可以提升到2 kHz左右, 使得配对样品的高通量表征成为了现实(见图21).这方面的应用实例包括: 生物膜和膜蛋白的静电性质(Medalsy & Müller 2013, Guo et al. 2016, Pfreundschuh et al. 2013), 被细菌排出的噬菌体(Alsteens et al. 2013a), 结合动物细胞表面受体的包膜病毒(Alsteens et al. 2017b), 结合人G蛋白偶联受体的配体(Alsteens et al. 2015), 两个配体与相同受体的结合(Pfreundschuh et al. 2015), 以及大型蛋白质复合物上配体结合事件的高清成像等(Pfreundschuh et al. 2014, Kim et al. 2015). ...
Quantitative imaging of the electrostatic field and potential generated by a transmembrane protein pore at subnanometer resolution.
1
2013
... 这些早期的实验主要受到设备采样率和压电陶瓷性能的限制, 其每条力曲线都需要接近数秒钟的时间完成, 这就大大制约了能够采集的力曲线的数量.由于每个像素至少要有一条力曲线才能计算机械性能, 所以早期的功能探针力谱结合成像技术结果往往像素点都比较少, 且实验时间比较长. 最近五年来, 随着设备性能的提升、新模式的推广, 以及探针相对样品运动轨迹的改良, 商品化设备的力曲线采集频率已经可以提升到2 kHz左右, 使得配对样品的高通量表征成为了现实(见图21).这方面的应用实例包括: 生物膜和膜蛋白的静电性质(Medalsy & Müller 2013, Guo et al. 2016, Pfreundschuh et al. 2013), 被细菌排出的噬菌体(Alsteens et al. 2013a), 结合动物细胞表面受体的包膜病毒(Alsteens et al. 2017b), 结合人G蛋白偶联受体的配体(Alsteens et al. 2015), 两个配体与相同受体的结合(Pfreundschuh et al. 2015), 以及大型蛋白质复合物上配体结合事件的高清成像等(Pfreundschuh et al. 2014, Kim et al. 2015). ...
Nanoscale three-dimensional patterning of molecular resists by scanning probes.
1
2010
... 除此以外, 通过基于力谱技术的原子力显微镜设备, 可以精确控制悬臂梁和探针在样品上移动轨迹和所施加的作用力.再加上可以选择具有不同物理性质的探针, 如镀有铂、金、钛等导电层或者镀有类金刚石耐磨层, 辅以施加在探针或样品上的电压, 还可以进行刻蚀和操纵实验等(Pires et al. 2010). 既可以应用于如图14所示的金属或半导体基底, 也可以用于生物样品表面的刻蚀和特定结构刻画(Liu et al. 2004, Paul et al.2011). 对基于原子力显微镜的力学操纵和刻蚀技术, Garcia等(2014)的综述文献中进行了详细报道. ...
Imaging articular cartilage tissue using atomic force microscopy (AFM).
1
2010
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
a. Nanomechanics of HaloTag tethers.
1
2013
... 力谱技术被用于研究特定力作用下样品的响应, 比如以较高的外力将蛋白质拉伸并去折叠后, 可以线性的放松作用力来观测其坍缩机制(Walther et al. 2007).观察结果表明各种蛋白质的坍缩过程中存在显著差异, 实验证实这些差异主要是由于各蛋白质的疏水相互作用差异产生的.该机制对于高度延展的蛋白质是常见的, 包括弹性蛋白质如PEVK.观察结果解释了高度延展的蛋白质的折叠行为与化学或热变性后折叠的差异.在HaloTag蛋白质机械性能研究的力钳实验中, 显示出HaloTag蛋白质对外力拉伸的灵敏度是L蛋白质的两倍, 并且再折叠的速率更低.实验展示了如何能够在高强度下长期观察蛋白质的折叠--去折叠循环, 而不会受到HaloTag栓系(tether)的干扰(Popa et al. 2013a). ...
b. Force dependency of biochemical reactions measured by single-molecule force-clamp spectroscopy.
... 将各种小分子修饰于针尖后研究其与配对目标的相互作用. (a)葡萄糖(Puntheeranurak et al. 2007), (b) 抗癌多肽(Shan et al. 2012), (c) MFZ2-12分子(Wildling et al. 2012), (d)聚环氧乙烷(Sonnenberg et al. 2007b) ...
Single-molecule reconstruction of oligonucleotide secondary structure by atomic force microscopy.
2
2014
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
... 原子力显微镜用于各尺寸生物样品成像示例. (a)核酸(Pyne et al. 2014); (b)蛋白质(Seelert et al. 2000); (c)纤维(Stamov et al. 2015); (d)病毒(Kuznetsov & McPherson 2011); (e)细胞(Friedrichs et al. 2007); (f)秀丽线虫横截面, 扫描范围24 \(\mu \)m \(\times \) 24 \(\mu \)m \(\times\) 6um (NT-MDT公司) ...
The interactions of spore coat morphogenetic proteins studied by single molecule recognition force spectroscopy. Chemistry -
Spatiotemporal dynamics of the nuclear pore complex transport barrier resolved by high-speed atomic force microscopy.
1
2016
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
An overview of the biophysical applications of atomic forcemicroscopy.
1
2004
... 原子力显微镜系统框图(Santosa et al. 2004) ...
One b hairpin after the other: exploring mechanical unfolding pathways of the transmembrane b-barrel protein OmpG.
The unfolding kinetics of ubiquitin captured with single-molecule force-clamp techniques.
1
2004
... 生物大分子在生理条件下(酸碱度近中性的室温稀盐水溶液)往往会形成规整、精确的超分子结构, 比如双链DNA 以双螺旋的形式存在, 蛋白质则经由\(\alpha \)-螺旋、\(\beta\)-折叠等结构完成组装. 为了研究生物大分子与水的相互作用, Cui等利用单分子力谱迫使高分子单链进入到不良溶剂中, 开展了生物大分子在水溶液与非极性溶剂中的对照研究(Cui et al. 2006, 2007). 双链DNA 在受到外力拉伸时, 可在约65 pN处观察到一个长平台, 对应于从B型双链DNA到其过度拉伸态的结构转变. 然而实验中发现, 在不同溶剂中力曲线存在着明显的变化.如果将水溶液替换为非极性溶剂(如二乙苯等), 不仅无法观测到指纹特征, 而且所得力曲线与单链DNA的一致.这一结果表明双链DNA在非极性溶剂中将变性为单链结构.在实验结果的基础上, 提出ssDNA与水分子之间的弱氢键可能是双链DNA稳定存在于水中的前提条件.原子力显微镜也被广泛应用与除DNA之外的许多小分子的力谱研究, 甚至包括了金颗粒(Xue et al. 2014)、纳米线(Marszalek et al. 2000)等非生物样品, 如图7(b)所示.早期的研究应用实例包括应用力谱分析技术在单分子水平上鉴定多糖混合物的组分(Marszalek et al.2001), 观察黄原胶分子与基底的结合力(Li et al. 1999a), 研究蚕丝素分子的纳米力学特性(Zhang et al. 2000), 观察肝素分子在外力作用下的力学性质(Marszalek et al. 2003), Fern\'{a}ndez课题组对泛素分子力学性质、及其与水分子相互作用的一系列研究等(Carrion-Vazquez et al.2003, Schlierf & Fern\'{a}ndez 2004, Brujić et al. 2006, Li et al. 2010). ...
Single-molecule unfolding force distributions reveal a funnel-shaped energy landscape.
1
2006
... 此后单个蛋白质去折叠过程的研究蓬勃发展, 对肌联蛋白展开的后续研究发现了其去折叠过程的中间态, 并描绘了其动力学能谱(Marszalek et al. 1999, Schlierf & Rief 2006), Li 等(2002)在此基础上将Titin 蛋白质分子作为一个弹簧处理, 从总体上了解其机械特性.笔者的实验进一步证明了去折叠过程中间态由于其与外力加载速率高度独立的性质, 起到了"力学缓冲"的作用, 其建立的动力学能量壁垒大大延长了I27蛋白质的寿命.实验数据建立了肌联蛋白动力学和拓扑结构间的联系.诸如I27的肌联蛋白末端结构域能够缓冲外力, 分子动力学模拟显示连接\(\beta \)折叠A的氢键断裂时形成了去折叠中间态.末端Ig蛋白质中, 连接\(\beta \)折叠A, B, G的氢键相互作用力, 能够在生理环境所能施加的外力加载速率下, 维持Ig蛋白质折叠结构能垒的稳定. 因此, 能够使蛋白质进入去折叠过程的作用力保持恒定.这一末端Ig中的机制能够使结构域在生理环境产生的外力下保持稳定, 从而在宏观上使肌联蛋白具有了外力吸收能力, 显著降低了意外进入去折叠过程的几率(Nunes et al. 2010).随着设备的进步, 目前已经可以在接近分子动态模拟的时间尺度上进行快速的去折叠实验(Rico et al. 2013). ...
Mapping mechanical force propagation through biomolecular complexes.
1
2015
... 膜蛋白去折叠过程会导致力曲线上产生很多锯齿状的突变峰, 每个突变峰均对应膜蛋白一个片段的去折叠过程.基于Bell-Evans模型推导出了蛋白质去折叠所需力与拉伸速率之间的关系, 揭示了蛋白质力学强度的动力学特性. 当拉伸速率较低时, 去折叠力将正比于拉伸速率, 对于较大的拉伸速率, 去折叠力与拉伸速率成指数关系(Bell 1978, Evans & Ritchie 1997). 通常利用虫链模型(worm-like chain)对力曲线上的各个突变峰进行拟合, 在虫链模型WLC拟合所得到的4个参数中, 每个结构域的能量势垒\(\DeltaG_{0}^\ddag \)的大小反映了跨越该能量势垒的难度, 此值越大越需要更大的作用力去克服, 可以反映结构域的稳定性; 活化态到结合态的距离\(x_{\rm u}\)反映了结构域能够容纳的构象变化, 此值越大则代表在去折叠过程中结构域可能出现的构象形态越多; 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\), 此值越大说明在自然条件下发生解离的速率越大, 结构域不稳定性越高; 等效弹性系数\(\kappa \)反映了结构域的机械力学参数, 此值越大表示结构域的机械硬度越强.通过上述参数可以描绘出去折叠动态过程的能量图谱(Janovjak et al. 2004, Kuo et al. 2010, Schoeler et al. 2015).也有研究表明这一能量图谱的表面粗糙而非平滑(Janovjak et al. 2007).近年来随着模型的优化已经有高度集成化的分析软件(Lamour et al. 2014), 同时随着设备性能的提升, 已经逐渐展开了这一模型在高速拉伸下的分析(Junker & Rief 2010, Berkovich et al. 2012). ...
Proton-powered turbine of a plant motor.
2
2000
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
... 原子力显微镜用于各尺寸生物样品成像示例. (a)核酸(Pyne et al. 2014); (b)蛋白质(Seelert et al. 2000); (c)纤维(Stamov et al. 2015); (d)病毒(Kuznetsov & McPherson 2011); (e)细胞(Friedrichs et al. 2007); (f)秀丽线虫横截面, 扫描范围24 \(\mu \)m \(\times \) 24 \(\mu \)m \(\times\) 6um (NT-MDT公司) ...
YidC assists the stepwise and stochastic folding of membrane proteins.
... 将各种小分子修饰于针尖后研究其与配对目标的相互作用. (a)葡萄糖(Puntheeranurak et al. 2007), (b) 抗癌多肽(Shan et al. 2012), (c) MFZ2-12分子(Wildling et al. 2012), (d)聚环氧乙烷(Sonnenberg et al. 2007b) ...
Nanostructured self-assembly of inverted formin 2 (INF2) and F-actin-INF2 complexes revealed by atomic force microscopy.
1
2014
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Force measurements of bacterial adhesion on metals using a cell probe atomic force microscope.
Structural changes in bacteriorhodopsin in response to alternate illumination observed by high-speed atomic force microscopy.
2
2011
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
Adhesion of melanoma cells to the surfaces of microspheres studied by atomic force microscopy.
Molecular mechanistic origin of the toughness of natural adhesives fibres and composites.
1
1999
... 弹性蛋白质在生物力学系统中是一个重要的功能基本单元, 因为它们具有理想的弹性力学强度和抗性. 弹性蛋白质是天然黏合剂, 可以作为细胞黏附和肌肉蛋白质弹性的基础, 也可以作为具有极好机械性能的结构材料.随着对蛋白质力学性质了解的不断深入, 采用蛋白质人工构建新型材料的研究有了重要的进展(Guerette et al. 1996, Smith et al. 1999, Ardell & Andersen 2001, Becker et al. 2003, Elvin et al. 2005, Lyons et al. 2007).例如人们已经能够利用已知的蛛丝蛋白质的结构合成具有非常好的延展性和强度的人工材料(Wong et al.2012), 这种方法也为制造下一代力学稳定的生物材料指明了方向(Heim et al. 2009). ...
Direct mechanical measurements of the elasticity of single DNA-molecules by using magnetic beads.
The relation between load and penetration in the axisymmetric Boussinesq problem for a punch of arbitrary profile.
1
1965
... 在细胞力学性质的研究中, 探索细胞杨氏模量随环境参数的变化是非常重要的一类实验.杨氏模量是弹性材料在一定的弹性范围内, 正向应力与正向应变的比值, 它反映材料的刚度. 杨氏模量初期主要应用于物理学领域, 目前在医学和生物学领域也得到了广泛应用.细胞的杨氏模量的各种拟合模型, 主要包括赫兹模型(Johnson et al. 1987)、Sneddon模型(Sneddon et al. 1965)、DMT(Derjaguin-Müller-Toporov)模型(Derjaguin et al. 1975)以及JKR模型(Johnson et al. 1971). ...
Exploring the folding pattern of a polymer chain in a single crystal by combining single-molecule force spectroscopy and steered molecular dynamics simulations.
1
2013
... 此外, Sonnenberg等对聚丙烯酸单层进行了力谱实验, 观察其相对缓冲液pH值的函数, 以研究连接着基底与针尖两个表面的聚合物的黏附性质(Sonnenberg et al. 2007a). 该研究表明, 聚丙烯酸链和氮化硅探针针尖之间的相互作用力取决于聚合物单层的覆盖密度, 以及聚合物链的轮廓长度.聚合物桥连的稳定性由两个表面的黏合强度决定, 作为依赖性聚电解质单层, 这一强度可以通过使用缓冲液pH值来进行调节.Zhang课题组使用了单分子力谱技术结合受控分子动力学(steered molecular dynamics, SMD)模拟的组合, 研究聚氧乙烯(PEO)单晶中高分子链之间的相互作用, 以及聚氧乙烯(PEO)链在其单晶中的折叠样式, 如图7(c)所示(Song et al.2013, Liu et al. 2011). 研究结果表明, 将单个PEO链从其单晶中提拉出来的作用力大小约为40 pN, 在稀溶液中形成的晶体中的PEO链的折叠样式遵循就近排列的重入折叠模型.在从熔体获得的晶体中, 力曲线上出现的大幅度波动主要是由于折叠样式上大而无规律的环状结构引起的.Liang和Fern\'{a}ndez(2011)的工作研究了单分子亲核取代化学反应中机械能与热效应之间的关系.通过单分子力谱技术研究了TCEP, 1, 4-\(_{\rm DL}\)-DTT和氢硫化物阴离子, 对受机械拉伸作用的多肽二硫键的影响.在一系列精确控制的温度下记录双分子亲核取代(S\(_{\rm N}\)2)事件, 对力和温度引起的反应速率的影响的比较表明, 每增大50 pN的拉力等效于温度升高10 K. Nash和Gaub(2012)测量了单一共聚物分子与金表面的黏附特性, 如图7(a)所示.研究者发现共聚物在高离子浓度缓冲液中的脱水和坍缩, 导致保持分子桥连长度的分布显著降低.在高盐浓度下聚合物显示出较低的绝对解离力, 而协同作用增加了多链相互作用中每条链对应的解离力. 研究结果证实, 坍缩构象中的聚合物在金表面上占据的体积显著减少.这些结果证明在单分子水平下, 溶剂诱导的环境响应共聚物的坍缩能够以可控的方式调节表面黏附力和桥接长度的分布. ...
b. Quantitative single molecule measurements on the interaction forces of poly(L-glutamic acid) with calcite crystals.
1
2007
... 此外, Sonnenberg等对聚丙烯酸单层进行了力谱实验, 观察其相对缓冲液pH值的函数, 以研究连接着基底与针尖两个表面的聚合物的黏附性质(Sonnenberg et al. 2007a). 该研究表明, 聚丙烯酸链和氮化硅探针针尖之间的相互作用力取决于聚合物单层的覆盖密度, 以及聚合物链的轮廓长度.聚合物桥连的稳定性由两个表面的黏合强度决定, 作为依赖性聚电解质单层, 这一强度可以通过使用缓冲液pH值来进行调节.Zhang课题组使用了单分子力谱技术结合受控分子动力学(steered molecular dynamics, SMD)模拟的组合, 研究聚氧乙烯(PEO)单晶中高分子链之间的相互作用, 以及聚氧乙烯(PEO)链在其单晶中的折叠样式, 如图7(c)所示(Song et al.2013, Liu et al. 2011). 研究结果表明, 将单个PEO链从其单晶中提拉出来的作用力大小约为40 pN, 在稀溶液中形成的晶体中的PEO链的折叠样式遵循就近排列的重入折叠模型.在从熔体获得的晶体中, 力曲线上出现的大幅度波动主要是由于折叠样式上大而无规律的环状结构引起的.Liang和Fern\'{a}ndez(2011)的工作研究了单分子亲核取代化学反应中机械能与热效应之间的关系.通过单分子力谱技术研究了TCEP, 1, 4-\(_{\rm DL}\)-DTT和氢硫化物阴离子, 对受机械拉伸作用的多肽二硫键的影响.在一系列精确控制的温度下记录双分子亲核取代(S\(_{\rm N}\)2)事件, 对力和温度引起的反应速率的影响的比较表明, 每增大50 pN的拉力等效于温度升高10 K. Nash和Gaub(2012)测量了单一共聚物分子与金表面的黏附特性, 如图7(a)所示.研究者发现共聚物在高离子浓度缓冲液中的脱水和坍缩, 导致保持分子桥连长度的分布显著降低.在高盐浓度下聚合物显示出较低的绝对解离力, 而协同作用增加了多链相互作用中每条链对应的解离力. 研究结果证实, 坍缩构象中的聚合物在金表面上占据的体积显著减少.这些结果证明在单分子水平下, 溶剂诱导的环境响应共聚物的坍缩能够以可控的方式调节表面黏附力和桥接长度的分布. ...
a. Choose Sides: Differential Polymer Adhesion.
2
2007
... 将各种小分子修饰于针尖后研究其与配对目标的相互作用. (a)葡萄糖(Puntheeranurak et al. 2007), (b) 抗癌多肽(Shan et al. 2012), (c) MFZ2-12分子(Wildling et al. 2012), (d)聚环氧乙烷(Sonnenberg et al. 2007b) ...
Photothermal cantilever actuation for fast single-molecule force spectroscopy.
1
2009
... 随着研究领域的深入, 人们意识到测量不同分离速度(即加载速率)的断裂力提供了关于结合键的动力学性质的信息(Moy et al.1994).结合键的动力学模型分析可以用于估计配体--受体键的热力学和动力学性质(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).这些性质包括跨越折叠状态的能量势垒\(\Delta G_{0}^\ddag \); 活化态到结合态的距离\(x_{\rm u}\); 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\); 此外结合键的寿命也可以通过使键处于低恒定力并等待键的断裂来测量(Woodside & Block 2014, Perez-Jimenez et al. 2011).此恒定作用力可以通过原子力显微镜的反馈系统以力钳模式施加(Oberhauser et al.2001, Stahl et al. 2009), 也可以利用天然的细胞膜栓系Tether来实现(Krieg et al. 2008b). ...
Imaging collagen type I fibrillogenesis with high spatiotemporal resolution.
3
2015
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
... 原子力显微镜用于各尺寸生物样品成像示例. (a)核酸(Pyne et al. 2014); (b)蛋白质(Seelert et al. 2000); (c)纤维(Stamov et al. 2015); (d)病毒(Kuznetsov & McPherson 2011); (e)细胞(Friedrichs et al. 2007); (f)秀丽线虫横截面, 扫描范围24 \(\mu \)m \(\times \) 24 \(\mu \)m \(\times\) 6um (NT-MDT公司) ...
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
Hydrostatic pressure and the actomyosin cortex drive mitotic cell rounding.
High-speed atomic force microscopy reveals rotary catalysis of rotorless F1-ATPase.
1
2011
... 成像是原子力显微镜应用最多、最广的领域, 绝大多数使用者都会用到这一功能. 在三十年的发展历程中, 已经开发出了十数种原子力显微镜的成像模式. 成像过程中, 大部分情况下需要将探针降落到样品表面, 调整悬臂梁相对于样品的高度以避免针尖和样品间的作用力过大或过小.按照每个像素得到的高度信息扫描整个视野, 便可以得到一个纳米甚至亚纳米级别分辨率的样品形貌信息. 在生物体中, 样品的范围可以从组织片段(Plodinec et al. 2010)、细胞(Friedrichs et al. 2007)、病毒(Kuznetsov & McPherson 2011)、人工或天然膜片(Mari et al. 2011)、各类纤维(Stamov et al. 2015)、水溶性蛋白质(Ido et al. 2014)、膜蛋白(Seelert et al. 2000)、到核酸(Ido et al. 2013, Pyne et al. 2014)等十分广泛.原子力显微镜成像可以用来直接观察细胞的分子机制, 包括ATP合成酶(Uchihashi et al. 2011)、传导通道(Müller et al. 2002)、成孔蛋白(Müller & Engel 1999, Mari et al. 2010)、毒素(Czajkowsky et al. 2004)、质子泵(Shibata et al. 2011)、离子通道(Mari et al. 2011, Rangl et al. 2016)、纤维蛋白(Cisneros et al. 2006)、丝状蛋白(Lehto et al. 2003, Sharma et al. 2014)、细胞外基质蛋白质(Friedrichs et al. 2007)等, 还可以用来观察其生长和重组过程(Gudzenko & Franz 2015). 图18分别以各尺寸的生物样品为例, 展示了原子力显微镜用于生物样品的成像结果. ...
Wnt11 functions in gastrulation by controlling cell cohesion through Rab5c and E{\-}cadherin.
0
2005
Probing protein-protein interactions in real time.
1
2000
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
Quantitative changes in the elasticity and adhesive properties of Escherichia coli ZK1056 prey cells during predation by Bdellovibrio bacteriovorus 109J.
0
2008
Signatures of hydrophobic collapse in extended proteins captured with force spectroscopy.
1
2007
... 力谱技术被用于研究特定力作用下样品的响应, 比如以较高的外力将蛋白质拉伸并去折叠后, 可以线性的放松作用力来观测其坍缩机制(Walther et al. 2007).观察结果表明各种蛋白质的坍缩过程中存在显著差异, 实验证实这些差异主要是由于各蛋白质的疏水相互作用差异产生的.该机制对于高度延展的蛋白质是常见的, 包括弹性蛋白质如PEVK.观察结果解释了高度延展的蛋白质的折叠行为与化学或热变性后折叠的差异.在HaloTag蛋白质机械性能研究的力钳实验中, 显示出HaloTag蛋白质对外力拉伸的灵敏度是L蛋白质的两倍, 并且再折叠的速率更低.实验展示了如何能够在高强度下长期观察蛋白质的折叠--去折叠循环, 而不会受到HaloTag栓系(tether)的干扰(Popa et al. 2013a). ...
Imaging glycosylation.
1
2008
... 1994年Florin等通过牛血清白蛋白将亲和素分子连接到AFM 针尖, 在覆盖了生物素分子的琼脂糖珠上测量了单对生物素--亲和素之间的相互作用力(160 \(\pm\) 20) pN. Hinterdorfer等(1996)研究了蛋白抗体与抗原的相互作用力, 并改进了蛋白质的固定方法, 通过一段PEG高分子聚合物链将蛋白质与表面相连, 这样不仅增加了抗体抗原的空间自由度, 而且能从力--距离曲线图上将其特异性相互作用与针尖和基底的非特异性相互作用力区分开来, 成为目前固定蛋白质的常用方法.在覆盖有抗原分子(人血清白蛋白)的云母表面测量了单对抗体-抗原之间的相互作用力(244 \(\pm\) 22) pN.将提纯的蛋白质分子吸附到平整基底进行分子力测量的优点是消除了细胞表面(如细胞形貌, 细胞膜的柔软动态特性)对蛋白质分子的影响, 从而显著提高了信噪比, 并且可通过对蛋白质分子同时进行形貌成像和分子力测量来证明检测结果的准确性, 因而提高了实验结果的可信度(Wang H et al. 2008).通过将提纯的蛋白质分子吸附到基底, 研究人员测量了不同类型蛋白质分子之间的相互作用力, 如铁蛋白(ferritin) (Allen et al. 1997)、钙黏蛋白(cadherin)(Baumgartner et al. 2000)、路易斯寡糖(sLeX)-选择素(selectin)(Zhang X et al. 2004)、Strep-Tactin与绿色荧光蛋白等(Baumann et al. 2016). ...
Evaluating protein attraction and adhesion to biomaterials with the atomic force microscope.
1
2004
... 此方法的一般步骤是将一个可伸缩的线性分子连接于探针, 再将需固定的分子连接于线性分子, 这种方法使被连接的分子具有一定的空间自由度(Chtcheglova et al. 2007). 针尖化学修饰的方法有多种, 其中比较常用的修饰方法是商品化的硅或氮化硅探针经化学处理, 如食人鱼洗液(98% H\(_{2}\)SO\(_{4}\):30%H\(_{2}\)O\(_{2}\)体积比7:3)清洗, 使其表面暴露出大量硅羟基(---SiOH), 再对探针表面硅羟基进一步硅烷化、醚化或氢硅化等, 或者直接将镀金针尖进行巯基化处理. 其中, 探针的硅烷化基于探针表面活化的硅羟基与硅烷化试剂反应, 如3 -氨基丙基三乙氧基硅烷(3-aminopropyltriethoxysilane, APTES)、3 -氨基丙基二甲基甲氧硅烷(3-aminopropyldimethylmethoxysilane, APS)等, 在针尖表面形成含有所需化学基团的有机单分子层(Xue et al. 2014).探针的醚化是利用醇类(通常是乙醇胺)与探针表面活化的羟基(---OH)反应, 形成末端含有氨基(---NH\(_{2})\)的有机单分子层(Wildling et al. 2011).在探针的氢硅化中, 首先对探针表面用1%的氢氟酸(HF)处理形成氢化的硅表面, 然后利用氢化表面与烯烃加成反应, 通过Si---C共价键将有机分子连接到针尖表面(Buriak 2002).镀金层探针的化学修饰方法, 是镀金探针表面与烷硫醇的巯基通过金硫键之间的强非特异性相互作用, 形成稳定的自组装单分子层(Wang et al. 2004). ...
The unfolding and folding dynamics of TNfnALL probed by single molecule force-ramp spectroscopy.
High-speed atomic force microscopy reveals strongly polarized movement of clostridial collagenase along collagen fibrils.
1
2016
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid.
0
1953
Stability enhancement of an atomic force microscope for long-term force measurement including cantilever modification for whole cell deformation.
2
2012
... 结合力谱技术的原子力显微镜成像模式在扫描时会记录数十万条力曲线数据, 用以反映机械性能或样品间的相互作用.因此缩短单条力曲线数据的记录时间, 能够减少图像获取时间并动态记录生物学的过程.然而有些配体--受体键合或者化学相互作用在非常快的外力作用下, 是远离平衡态的破坏状态, 此时想要推导其平衡态的力学行为是比较困难的(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).因为在高加载速率下, 必须考虑悬臂梁的流体动力学阻力和物理限制(Amo & Garcia 2016). 所以想要反映这类目标接近平衡态时的相互作用, 需要缓慢地施加外力. 因此, 似乎有可能需要开发能够在非常大的范围选择加载速率的方法, 例如结合当前的超稳定AFM方向(King et al. 2009, Bull et al. 2014, Weafer et al. 2012)与超高速AFM方向(Dong et al. 2011, Rico et al. 2013). ...
... (4)AFM技术的进一步突破还依赖于稳定激光系统的研制、悬臂梁系统的制作、压电换能器材料的探索、光电检测器的改进; 同时也与电子线路、传动马达系统、隔震隔噪声系统的设计与完善密切相关.由于原子力显微镜分辨率和功能与探针性能的密切相关性, 因而探针的制造工艺的提高也是亟待解决的关键问题.目前已经有研究组通过控制温度、悬臂镀层等一系列方法, 来提高AFM测量力值的长期稳定性, 使得AFM 针尖的漂移显著降低(Weafer et al.2012). ...
The fuzzy coat of pathological human Tau fibrils is a two-layered polyelectrolyte brush.
1
2013
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
... 将各种小分子修饰于针尖后研究其与配对目标的相互作用. (a)葡萄糖(Puntheeranurak et al. 2007), (b) 抗癌多肽(Shan et al. 2012), (c) MFZ2-12分子(Wildling et al. 2012), (d)聚环氧乙烷(Sonnenberg et al. 2007b) ...
Linking of sensor molecules with amino groups to amino-functionalized AFM tips.
1
2011
... 此方法的一般步骤是将一个可伸缩的线性分子连接于探针, 再将需固定的分子连接于线性分子, 这种方法使被连接的分子具有一定的空间自由度(Chtcheglova et al. 2007). 针尖化学修饰的方法有多种, 其中比较常用的修饰方法是商品化的硅或氮化硅探针经化学处理, 如食人鱼洗液(98% H\(_{2}\)SO\(_{4}\):30%H\(_{2}\)O\(_{2}\)体积比7:3)清洗, 使其表面暴露出大量硅羟基(---SiOH), 再对探针表面硅羟基进一步硅烷化、醚化或氢硅化等, 或者直接将镀金针尖进行巯基化处理. 其中, 探针的硅烷化基于探针表面活化的硅羟基与硅烷化试剂反应, 如3 -氨基丙基三乙氧基硅烷(3-aminopropyltriethoxysilane, APTES)、3 -氨基丙基二甲基甲氧硅烷(3-aminopropyldimethylmethoxysilane, APS)等, 在针尖表面形成含有所需化学基团的有机单分子层(Xue et al. 2014).探针的醚化是利用醇类(通常是乙醇胺)与探针表面活化的羟基(---OH)反应, 形成末端含有氨基(---NH\(_{2})\)的有机单分子层(Wildling et al. 2011).在探针的氢硅化中, 首先对探针表面用1%的氢氟酸(HF)处理形成氢化的硅表面, 然后利用氢化表面与烯烃加成反应, 通过Si---C共价键将有机分子连接到针尖表面(Buriak 2002).镀金层探针的化学修饰方法, 是镀金探针表面与烷硫醇的巯基通过金硫键之间的强非特异性相互作用, 形成稳定的自组装单分子层(Wang et al. 2004). ...
Materials by design: Merging proteins and music.
1
2012
... 弹性蛋白质在生物力学系统中是一个重要的功能基本单元, 因为它们具有理想的弹性力学强度和抗性. 弹性蛋白质是天然黏合剂, 可以作为细胞黏附和肌肉蛋白质弹性的基础, 也可以作为具有极好机械性能的结构材料.随着对蛋白质力学性质了解的不断深入, 采用蛋白质人工构建新型材料的研究有了重要的进展(Guerette et al. 1996, Smith et al. 1999, Ardell & Andersen 2001, Becker et al. 2003, Elvin et al. 2005, Lyons et al. 2007).例如人们已经能够利用已知的蛛丝蛋白质的结构合成具有非常好的延展性和强度的人工材料(Wong et al.2012), 这种方法也为制造下一代力学稳定的生物材料指明了方向(Heim et al. 2009). ...
Reconstructing folding energy landscapes by single-molecule force spectroscopy.
1
2014
... 随着研究领域的深入, 人们意识到测量不同分离速度(即加载速率)的断裂力提供了关于结合键的动力学性质的信息(Moy et al.1994).结合键的动力学模型分析可以用于估计配体--受体键的热力学和动力学性质(Evans & Calderwood 2007, Dudko et al. 2008, Friddle et al. 2012).这些性质包括跨越折叠状态的能量势垒\(\Delta G_{0}^\ddag \); 活化态到结合态的距离\(x_{\rm u}\); 不施加外力平衡态条件下的复合物解离速率常数\(k_{\rm off}(0)\); 此外结合键的寿命也可以通过使键处于低恒定力并等待键的断裂来测量(Woodside & Block 2014, Perez-Jimenez et al. 2011).此恒定作用力可以通过原子力显微镜的反馈系统以力钳模式施加(Oberhauser et al.2001, Stahl et al. 2009), 也可以利用天然的细胞膜栓系Tether来实现(Krieg et al. 2008b). ...
Quantifying thiol-gold interactions towards the efficient strength control.
2
2014
... 此方法的一般步骤是将一个可伸缩的线性分子连接于探针, 再将需固定的分子连接于线性分子, 这种方法使被连接的分子具有一定的空间自由度(Chtcheglova et al. 2007). 针尖化学修饰的方法有多种, 其中比较常用的修饰方法是商品化的硅或氮化硅探针经化学处理, 如食人鱼洗液(98% H\(_{2}\)SO\(_{4}\):30%H\(_{2}\)O\(_{2}\)体积比7:3)清洗, 使其表面暴露出大量硅羟基(---SiOH), 再对探针表面硅羟基进一步硅烷化、醚化或氢硅化等, 或者直接将镀金针尖进行巯基化处理. 其中, 探针的硅烷化基于探针表面活化的硅羟基与硅烷化试剂反应, 如3 -氨基丙基三乙氧基硅烷(3-aminopropyltriethoxysilane, APTES)、3 -氨基丙基二甲基甲氧硅烷(3-aminopropyldimethylmethoxysilane, APS)等, 在针尖表面形成含有所需化学基团的有机单分子层(Xue et al. 2014).探针的醚化是利用醇类(通常是乙醇胺)与探针表面活化的羟基(---OH)反应, 形成末端含有氨基(---NH\(_{2})\)的有机单分子层(Wildling et al. 2011).在探针的氢硅化中, 首先对探针表面用1%的氢氟酸(HF)处理形成氢化的硅表面, 然后利用氢化表面与烯烃加成反应, 通过Si---C共价键将有机分子连接到针尖表面(Buriak 2002).镀金层探针的化学修饰方法, 是镀金探针表面与烷硫醇的巯基通过金硫键之间的强非特异性相互作用, 形成稳定的自组装单分子层(Wang et al. 2004). ...
... 生物大分子在生理条件下(酸碱度近中性的室温稀盐水溶液)往往会形成规整、精确的超分子结构, 比如双链DNA 以双螺旋的形式存在, 蛋白质则经由\(\alpha \)-螺旋、\(\beta\)-折叠等结构完成组装. 为了研究生物大分子与水的相互作用, Cui等利用单分子力谱迫使高分子单链进入到不良溶剂中, 开展了生物大分子在水溶液与非极性溶剂中的对照研究(Cui et al. 2006, 2007). 双链DNA 在受到外力拉伸时, 可在约65 pN处观察到一个长平台, 对应于从B型双链DNA到其过度拉伸态的结构转变. 然而实验中发现, 在不同溶剂中力曲线存在着明显的变化.如果将水溶液替换为非极性溶剂(如二乙苯等), 不仅无法观测到指纹特征, 而且所得力曲线与单链DNA的一致.这一结果表明双链DNA在非极性溶剂中将变性为单链结构.在实验结果的基础上, 提出ssDNA与水分子之间的弱氢键可能是双链DNA稳定存在于水中的前提条件.原子力显微镜也被广泛应用与除DNA之外的许多小分子的力谱研究, 甚至包括了金颗粒(Xue et al. 2014)、纳米线(Marszalek et al. 2000)等非生物样品, 如图7(b)所示.早期的研究应用实例包括应用力谱分析技术在单分子水平上鉴定多糖混合物的组分(Marszalek et al.2001), 观察黄原胶分子与基底的结合力(Li et al. 1999a), 研究蚕丝素分子的纳米力学特性(Zhang et al. 2000), 观察肝素分子在外力作用下的力学性质(Marszalek et al. 2003), Fern\'{a}ndez课题组对泛素分子力学性质、及其与水分子相互作用的一系列研究等(Carrion-Vazquez et al.2003, Schlierf & Fern\'{a}ndez 2004, Brujić et al. 2006, Li et al. 2010). ...
Single-molecule imaging on living bacterial cell surface by high-speed AFM.
1
2012
... 30年来, 原子力显微镜设备本身, 在技术以及方法上也有了重大的革新.很长一段时间内AFM成像中的时间分辨率是相当低的, 限制了其跟踪动态过程的能力. 如今, 关键技术的进步使成像速度得到了显著提高, 衍生出了称为高速AFM的操作模式(Hansma et al. 2006, Ando et al. 2013, Adams et al. 2016).这些进展包括引入具有超短响应时间的小悬臂、更好地抑制机械振动(Sullan et al.2013)、开发快速稳定的压电扫描器, 以及使用在成像期间动态调节反馈增益的控制器等. 如今, 这些改进使得有可能在生理环境中直接观察一系列生理反应的动态过程, 这其中包括分子伴侣(Viani et al. 2000), 细胞骨架运动蛋白(Kodera et al. 2010), 光驱质子泵(Shibata et al. 2011)和酶旋转马达(Uchihashi et al.2011), 胶原纤维发生(Stamov et al. 2015)和酶促降解(Watanabe-Nakayama et al. 2016), 蛋白质在脂膜层中的组装(Karner et al. 2016, Chiaruttini et al. 2015), 蛋白质在活细菌膜层中的组装(Yamashita et al. 2012), 以及核孔复合物的时空动力学(Sakiyama et al. 2016)等. ...
a. Energy landscape of aptamer/protein complexes studied by single-molecule force spectroscopy. Chemistry -
Coexistence of ribbon and helical fibrils originating from hIAPP(20-29) revealed by quantitative nanomechanical atomic force microscopy.
1
2013
... 原子力显微镜用于各尺寸生物样品成像示例(Dufrêne et al. 2013). (a)探针像对于样品不断进行扫描的同时, 还在往复运动记录力曲线; (b)每个像素点上至少会得到一条力曲线, 通过其直接计算力学相关参数; (c)膜蛋白的形貌图、杨氏模量图、样品形变量图、黏附力图, 标尺为200 nm基于力谱技术的成像已经被应用到多个类型的生物样品, 包括分裂动物细胞的沟槽变硬(Matzke et al. 2001), 各种动物和细菌细胞(Heu et al. 2012, Hecht et al. 2015, Formosa-Dague et al. 2016, Beaussart et al. 2015), 病毒(Marchetti et al.2016), 生物膜和膜蛋白(Medalsy & Müller 2013, Dong et al. 2009, Medalsy et al. 2011).和淀粉样蛋白原纤维(Wegmann et al.2013, Zhang S et al. 2013)等. ...
Quantitative biomolecular imaging by dynamic nanomechanical mapping.
1
2014
... 近年来, 力谱技术与原子力成像模式的组合应用, 使生物样品的多参数成像成为可能(Zhang S et al. 2014).这种组合通常被不同的原子力显微镜厂商命名为不同的模式名称, 各厂商之间的模式一般会在针尖相对样品的运动轨迹, 或者力曲线数据采集和拟合的方式之间存在着细微的差异.但是这些都是基于力--距离曲线, 即原子力显微镜力谱技术的成像模式.这种成像模式的工作原理如图19所示, 针尖在样品表面不断地重复接触-回退的力曲线轨迹, 每个像素点至少记录一条力曲线, 从而通过力曲线直接提取各种机械参数.这些参数中包括样品的弹性、刚度、形变量, 针尖的能量耗散等一系列指标. ...
Quantification of the lateral detachment force for bacterial cells using atomic force microscope and centrifugation.
EMSA and single-molecule force spectroscopy study of interactions between bacillus subtilis single-stranded DNA-binding protein and single-stranded DNA.
0
2011
Single-molecule force spectroscopy on bombyx mori silk fibroin by atomic force microscopy.
1
2000
... 生物大分子在生理条件下(酸碱度近中性的室温稀盐水溶液)往往会形成规整、精确的超分子结构, 比如双链DNA 以双螺旋的形式存在, 蛋白质则经由\(\alpha \)-螺旋、\(\beta\)-折叠等结构完成组装. 为了研究生物大分子与水的相互作用, Cui等利用单分子力谱迫使高分子单链进入到不良溶剂中, 开展了生物大分子在水溶液与非极性溶剂中的对照研究(Cui et al. 2006, 2007). 双链DNA 在受到外力拉伸时, 可在约65 pN处观察到一个长平台, 对应于从B型双链DNA到其过度拉伸态的结构转变. 然而实验中发现, 在不同溶剂中力曲线存在着明显的变化.如果将水溶液替换为非极性溶剂(如二乙苯等), 不仅无法观测到指纹特征, 而且所得力曲线与单链DNA的一致.这一结果表明双链DNA在非极性溶剂中将变性为单链结构.在实验结果的基础上, 提出ssDNA与水分子之间的弱氢键可能是双链DNA稳定存在于水中的前提条件.原子力显微镜也被广泛应用与除DNA之外的许多小分子的力谱研究, 甚至包括了金颗粒(Xue et al. 2014)、纳米线(Marszalek et al. 2000)等非生物样品, 如图7(b)所示.早期的研究应用实例包括应用力谱分析技术在单分子水平上鉴定多糖混合物的组分(Marszalek et al.2001), 观察黄原胶分子与基底的结合力(Li et al. 1999a), 研究蚕丝素分子的纳米力学特性(Zhang et al. 2000), 观察肝素分子在外力作用下的力学性质(Marszalek et al. 2003), Fern\'{a}ndez课题组对泛素分子力学性质、及其与水分子相互作用的一系列研究等(Carrion-Vazquez et al.2003, Schlierf & Fern\'{a}ndez 2004, Brujić et al. 2006, Li et al. 2010). ...
Molecular basis of the dynamic strength of the sialyl Lewis X-selectin interaction.
1
2004
... 1994年Florin等通过牛血清白蛋白将亲和素分子连接到AFM 针尖, 在覆盖了生物素分子的琼脂糖珠上测量了单对生物素--亲和素之间的相互作用力(160 \(\pm\) 20) pN. Hinterdorfer等(1996)研究了蛋白抗体与抗原的相互作用力, 并改进了蛋白质的固定方法, 通过一段PEG高分子聚合物链将蛋白质与表面相连, 这样不仅增加了抗体抗原的空间自由度, 而且能从力--距离曲线图上将其特异性相互作用与针尖和基底的非特异性相互作用力区分开来, 成为目前固定蛋白质的常用方法.在覆盖有抗原分子(人血清白蛋白)的云母表面测量了单对抗体-抗原之间的相互作用力(244 \(\pm\) 22) pN.将提纯的蛋白质分子吸附到平整基底进行分子力测量的优点是消除了细胞表面(如细胞形貌, 细胞膜的柔软动态特性)对蛋白质分子的影响, 从而显著提高了信噪比, 并且可通过对蛋白质分子同时进行形貌成像和分子力测量来证明检测结果的准确性, 因而提高了实验结果的可信度(Wang H et al. 2008).通过将提纯的蛋白质分子吸附到基底, 研究人员测量了不同类型蛋白质分子之间的相互作用力, 如铁蛋白(ferritin) (Allen et al. 1997)、钙黏蛋白(cadherin)(Baumgartner et al. 2000)、路易斯寡糖(sLeX)-选择素(selectin)(Zhang X et al. 2004)、Strep-Tactin与绿色荧光蛋白等(Baumann et al. 2016). ...
Direct measurements of the interaction between pyrene and graphite in aqueous media by single molecule force spectroscopy: Understanding the pi-pi interactions.

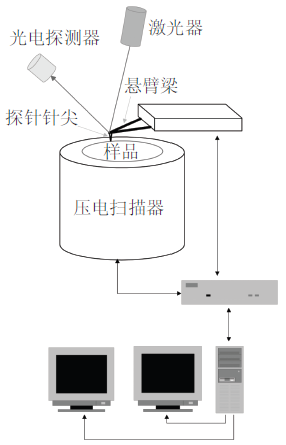
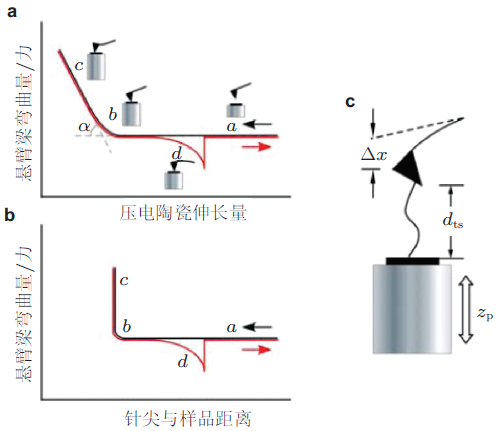
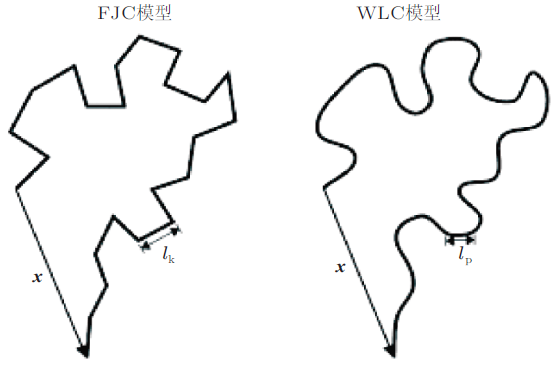






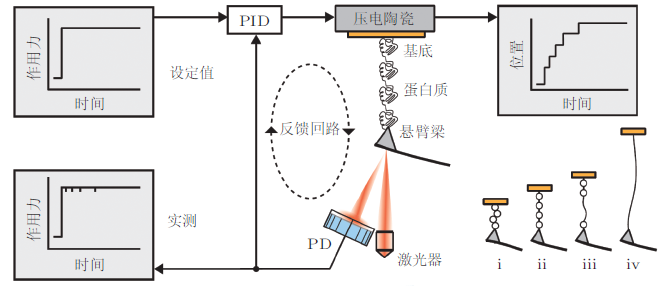
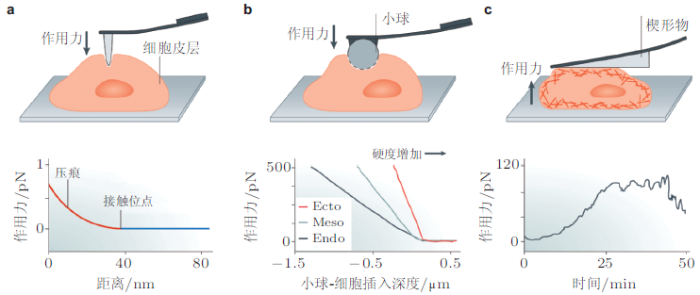
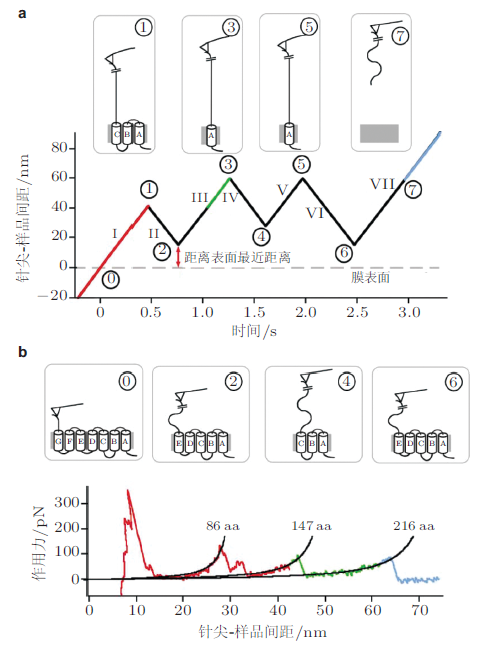

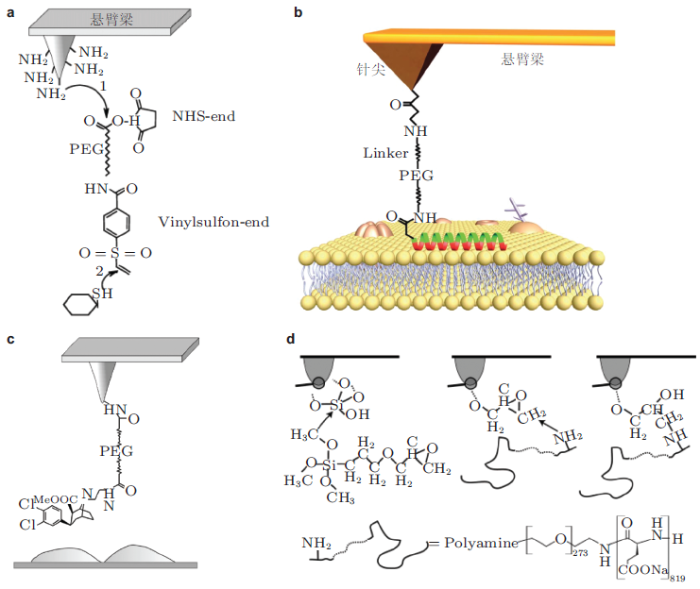
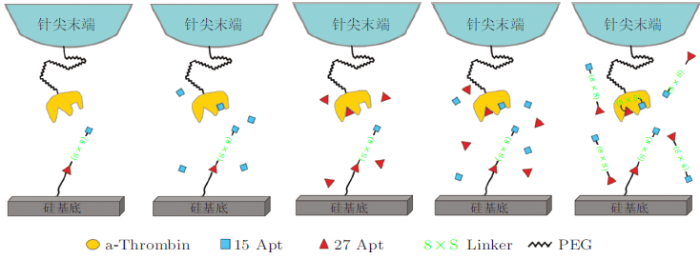
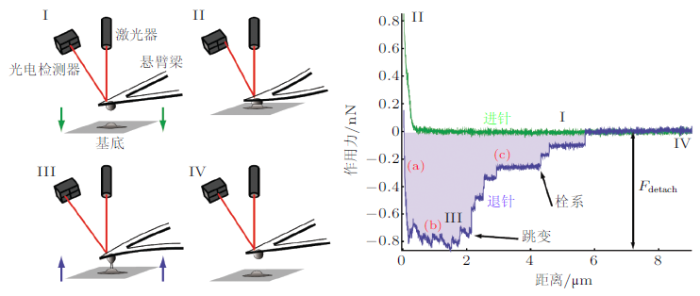
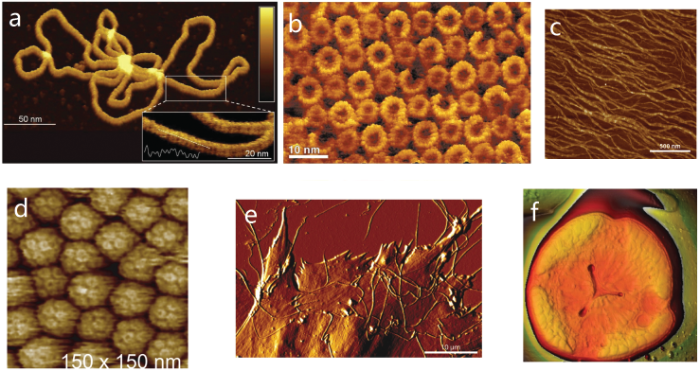

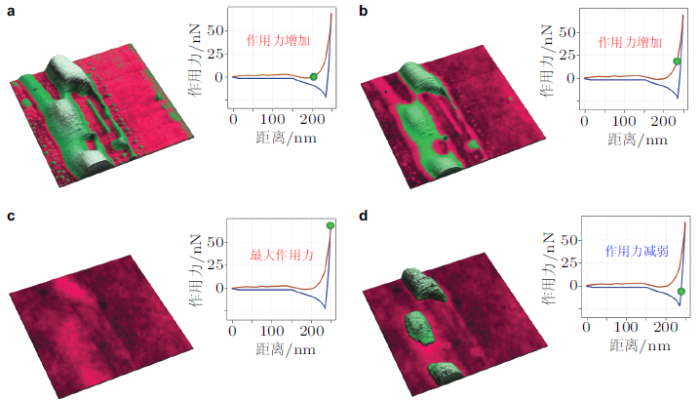

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
